FECUNDACION
La fecundación, es el proceso por el cual
dos gametos (masculino y femenino) se fusionan durante la reproducción sexual para crear un
nuevo individuo con un genoma derivado de ambos progenitores.
Los dos fines principales de la fecundación son la combinación de genesderivados de ambos progenitores y la generación de un nuevo individuo.
PROCESOS:
Los detalles de la
fecundación son tan diversos como las especies; sin embargo, existen cuatro
eventos que son constantes en todas ellas:
1.
El primer contacto y reconocimiento entre el óvulo y el espermatozoide, que en la mayor parte de los casos
es de gran importancia para asegurar que los gametos sean de la misma especie.
2.
La regulación de la interacción entre el espermatozoide y el gameto
femenino. Solamente un gameto masculino debe fecundar un gameto femenino. Esto
puede lograrse permitiendo que sólo un espermatozoide entre en el óvulo, lo que
impedirá el ingreso de otros.
3.
La fusión del material genético proveniente de ambos gametos.

RECONOCIMIENTO ENTRE GAMETO MASCULINO
Y FEMENINO
Se producen los
siguientes pasos:
1.
Quimio-atracción desde el gameto femenino hacia el masculino, por medio
de la secreción de moléculas solubles que atraen al espermatozoide. En un gran
número de especies los espermatozoides son atraídos hacia el gameto femenino, a
través de la secreción de una sustancia química por parte de este último.
2.
Exocitosis de la vesícula acrosómica del
gameto masculino de modo que se puedan liberar las enzimascontenidas en esta vesícula.
3.
Unión del gameto masculino a la capa extracelular que recubre el óvulo. El espermatozoide se une primero al
gameto femenino, ocurriendo después la liberación del contenido de la vesícula
acrosómica.
4.
Paso del espermatozoide a través de la matriz extracelular (membrana vitelina
en el erizo de mar; zona pelúcidaen mamíferos) que recubre al óvulo.
5.
Fusión de las membranas celulares del gameto femenino y masculino.
Motilidad de los espermatozoides
En distintas
regiones del tracto reproductivo femenino se secretan distintas moléculas, las cuales pueden influenciar
la motilidad de los
espermatozoides. Por ejemplo, en algunos casos de roedores, los espermatozoides al pasar del
útero a las tropas se Falopio se hiperactivan, nadando a velocidades mayores.
La hiperactivación parece estar relacionada con cAMP de un canal de calcio en
la cola de los espermatozoides. Esto facilita la motilidad a través de fluidos viscosos como los que se encuentran en
las trompas de Falopio. La hiperactividad y la hialuronidasa permite que los espermatozoides
atraviesen la capa del cúmulo.
Otros factores
secretados en el oviducto proporcionan
el componente direccional del movimiento de los espermatozoides, se especula
que estos factores quimiotácticos son secretados por el folículo ovárico.
También se ha visto que solo los espermatozoides capacitados logran ser
atraídos por el fluido folicular quimiotáctico.

Barreras a superar por los espermatozoides
La capacitación
permite al espermatozoide sobrepasar varias barreras y lograr la fecundación.
La primera barrera al llegar al huevo es una capa de células cúmulus en ácido
hialurónico. La actividad de la hialuronidasa en la superficie de la cabeza del
espermatozoide lo ayuda a penetrar esta barrera. La segunda barrera es la zona
pelúcida, la cual es una capa de glucoproteínas. El espermatozoide logra penetrar
esta barrera gracias a la reacción acrosómica (liberación de contenidos de la
vesícula acrosómica ubicada en la cabeza del espermatozoide).
La zona pelúcida
La zona pelúcida
juega un papel análogo a la membrana vitelina de los invertebrados. Esta matriz
extracelular, la cual es sintetizada por el ovocito tiene dos funciones
principales: unir el espermatozoide y comenzar la reacción acrosómica. Esta
capa de la zona pelúcida tiene tres glucoproteínas principales ZP1, ZP2 y ZP3.
Esta última es un receptor especie-específica para la unión del espermatozoide.
Unión del espermatozoide con el huevo
La membrana celular que recubre la cabeza del espermatozoide tiene
varias proteínas (un ejemplo es la SED1), estas proteínas se pueden unir
cientos de glucoproteínas de ZP3 de la zona pelúcida (por cadenas de
carbohidratos enlazadas por serina y treonina. Al experimentar la reacción
acrosómica sobre la zona pelúcida, los espermatozoides concentran sus enzimas
proteolíticas sobre el punto de adhesión y digieren un agujero a través de esta
capa extracelular. Cuando ZP3 se enlaza a los receptores en la membrana celular
del espermatozoide se activa la reacción acrosómica.
Una de las proteínas del espermatozoide enlazada es la
galactosiltranferasa-I, una enzima intramembranosa cuyos sitios activos miran
hacia el exterior y se une a los residuos de carbohidratos de la ZP3. Esto a la
vez activa las proteínas G específicas de la membrana del espermatozoide, las
cuales activan una cascada que abre los canales de calcio y provoca la
exocitosis del contenido acrosómico, esto está mediado por el calcio de la
vesícula acrosómica.
El contenido
acrosómico incluye β-N-acetilglucosaminidasa y varias proteasas las cuales
rompen cadenas de oligosacáridos de las glucoproteínas de la zona pelúcida.
Esto permite que el espermatozoide perfore la zona pelúcida y se acerque a la
membrana plasmática del huevo. Para la perforación continua sin perder la
adhesión con la matriz extracelular se debe logar una unión secundaria a la
zona pelúcida, a través de ZP2.
La reacción acrosómica también
expone unas proteínas de la superficie del espermatozoide que se pueden unir
con la membrana del huevo y las cuales permiten la fusión de ambas membranas.
Otro componente importante es la proteína fertilina la cual se une a un
receptor tipo integrina en la membrana. Un receptor del huevo para reconocer el
espermatozoide es la proteína CD9 la cual inicia la integración
huevo-espermatozoide y que luego es un factor crítico en la fusión de los
miocitos (precursores musculares) para formar el miotubo multinucleado del
músculo estriado. En mamíferos el espermatozoide no contacta al gameto femenino
en su extremo, sino sobre el costado de la cabeza, en la región dominio
ecuatorial de la cabeza del espermatozoide.
Para evitar la
polispermia, apenas llega el primer espermatozoide a la membrana plasmática del
huevo y comienza la integración, se liberan los gránulos corticales los cuales
contienen enzimas que evitan la unión de otros espermatozoides con la zona
pelúcida. A diferencia de otros organismos como los erizos de mar, en mamíferos
no se presenta un cambio de potencial de membrana.

Activación del huevo a partir de la fertilización
La activación del
huevo a partir de la fertilización activa una serie de eventos que resultan en
el comienzo del desarrollo. Los eventos principales son: el huevo completa
meiosis, se unen los núcleos del huevo y el espermatozoide para formar un
cigoto diploide, y el huevo fertilizado entra a mitosis. En el caso de ratones
y humanos las membranas de los pronúcleos desaparecen antes de la unión de
estos.
Al igual que en
erizos de mar la activación del huevo está relacionado con la liberación de
iones libres de calcio en el huevo (produciendo una ola de calcio la cual es necesaria
y suficiente para comenzar el desarrollo). La ola de calcio comienza en el
punto por el cual entró el espermatozoide y cruza todo el huevo. Hay
oscilaciones en la concentración de calcio por varias horas después de la
fertilización.
El mecanismo por el cual se inicia la
liberación de calcio no es conocido, pero se cree que el espermatozoide induce
factores específicos de proteína que inicia la liberación de calcio después de
la fusión de los gránulos corticales. Al aumentar las concentraciones de calcio
se inicia el desarrollo del huevo fertilizado al activar proteínas relacionadas
con el ciclo celular.

DIVISION DE LOS BLASTOMEROS
Las blastómeras son
un tipo de células embrionarias animales indiferenciadas resultantes de
la segmentación del cigotodespués de la fecundación.
En el ser humano, las blastómeras son producidas
después de la fecundación y se dividen por Mitosis. Dos días después de la fecundación
hay 4 blastómeros, y entre 7 y 8 al tercer día.
MORULA
Las blastómeras se dividen rápida y repetidamente durante los primeros
días de la gestación aunque el tamaño del embrión permanece constante (aumenta el
número de células pero no su tamaño). Pasados cuatro días, el huevo fecundado
tiene más de 32 blastómeros, recibiendo a partir de ese momento el nombre
de mórula. Posteriormente se produce la blastulación, que representa el inicio de la diferenciación celular de tal manera que un grupo de blastómeros
se sitúan formando una envoltura externa denominada trofoblasto, que posteriormente dará origen a la placenta,
mientras que otros se agrupan constituyendo la masa celular interna o embrioblasto que dará origen al embrión.
BLASTOCELE
El blastocele es una cavidad llena de líquido y que junto al trofoectodermo y la
masa celular interna forman el blastocisto.

IMPLANTACION
La implantación del embrión humano es
el proceso por el que el (cigoto) se ancla al endometrio. Es una de las fases de la embriogénesis humana.
La implantación
comienza al final de la primera semana —séptimo u octavo día— después de
la fecundación del óvulo por el espermatozoide y se extiende hasta el final de
la segunda semana —14 días después de la fecundación—.
El óvulo fecundado
por el espermatozoide forma el cigoto que se dividirá convirtiéndose en
la mórula y después en
el blastocisto o blástula que se adherirá a la pared
del útero.
El conjunto de
células que ha formado el cigoto, pasa por la fase morula y de blastocisto o blástula. El blastocisto se encuentra
dividido en dos grupos de células; uno, más externo, y otro más interno. El
grupo interno, se convertirá en el embrión, y el exterior, en la membrana que
lo protegerá y nutrirá durante el embarazo.
La implantación o
adherencia al útero permite que el embrión reciba oxígeno y nutrientes de la madre —a
través de la sangre— para su desarrollo y crecimiento. El embrión adherido a la
pared del útero comienza a desarrollarse, para lo cual emite unas
prolongaciones arborescentes hacia la mucosa uterina que le permite adherirse
al útero y así extraer los nutrientes necesarios de la madre.

PROCESO DE
IMPLANTACION
El proceso de implantación se divide en dos periodos: un
período preimplantatorio, durante el cual ocurre la preparación del endometrio,
la preparación del ovocito y la aposición; y un período implantatorio que a su
vez se divide en adhesión, ruptura de la barrera epitelial e invasión.
Preparación del endometrio:
La producción de estrógeno
y progesterona es esencial para el proceso de implantación. Ambas
hormonas están implicadas en numerosas cascadas de señalización autocrinas y
paracrinas que van a desencadenar la fijación y posterior invasión del embrión
en el útero materno.
Durante la fase
folicular, el endometrio experimenta una fase mitogénica y se desarrolla
hasta alcanzar un espesor de entre 8-12 mm. Esta fase está mediada por el
estradiol (estrógeno) que media la expresión de numerosas citocinas y factores
de crecimiento y también, de los receptores de progesterona.
Tras la ovulación,
el cuerpo lúteo actúa como una glándula endocrina secretando progesterona que
va a inducir la expresión de otra serie de citocinas y factores responsables de
los cambios que sufre el endometrio para la implantación, además de inhibir la
producción de receptores de estrógenos y favorecer la síntesis de
17-β-hidroxilasadeshidrogenasa, que transforma el estradiol en una forma menos
activa. Los principales cambios que hacen receptivo al endometrio son:
·
Disminución de la fase mitogénica
·
Formación de pinópodos
·
Decidualización del endometrio
·
Disminución de las uniones estrechas o
“tightjuntion” entre las células epiteliales que van a facilitar la
invasión del embrión al epitelio
·
Apoptosis regulada localmente que
también facilitará la invasión (es objeto de estudios actualmente)
Predecidulación y decidulación del
endometrio:
La predecidulación ocurre aproximadamente 7 días
después de la ovulación y consiste en el aumento de grosor del tejido
endometrial, aumento de la vascularización y crecimiento de las glándulas para
potenciar las secreciones. Hacia el noveno o décimo día tras la ovulación, las
células de la superficie del endometrio se constituyen en una capa de células
redondeadas denominadas deciduales que desaparece si no ocurre el embarazo, al
igual que las glándulas que se atrofiarán y degenerarán a menos que ocurra la
implantación. La decidulación es
una etapa de expansión de la explicada anteriormente que tiene lugar unas 24
horas antes de la aposición del blastocito en el endometrio. Se favorece el
desarrollo de las glándulas, las células deciduales almacenan lípidos y
glucógeno en grandes cantidades y toman una forma poliédrica, hay una
reorganización vascular aumentando la permeabilidad en el sitio donde ocurrirá
la implantación. La capa de células deciduales o decidua permanecerá durante el
primer trimestre del embarazo al menos aunque su función como tejido
circundante es sustituida por la placenta. Es probable que esta etapa sea
promovida y mantenida por el propio blastocisto ya que existe un mayor grado de
decidulación en los ciclos con concepción o cuando se simula la invasión
natural del embrión que en aquellos en los que no ocurre.
FORMACION DE PINOPODOS:
La formación de pinópodos, pequeñas protuberancias con forma
digital que solo están presentes en el periodo de ventana de implantación
promovida por la progesterona. Los pinópodos absorben parte del fluido
(pinocitosis) y macromoléculas (endocitosis) presentes en la cavidad uterina,
lo que acerca al blastocito al endometrio y favorece el contacto entre ambos.
El endometrio también debe secretar ciertos nutrientes para abastecer al
embrión las 72 horas que pasa en la cavidad uterina sin implantarse. Y también
secreta varias proteínas dependientes de esteroides y sustancias de la matriz
(moléculas de adhesión, receptores de las moléculas de la matriz) que facilitan
la implantación.

PREPARACION DEL EMBRION:
La eclosión del blastocito es imprescindible para que ocurra la
implantación. Este proceso ocurre seis días tras la fecundación y consiste en
la roture de la zona pelúcida (ZP) que se encuentra limitando el embrión.
Intervienen factores líticos uterinos y factores del propio blastocito. Un buen
candidato es la plasmina, molécula con actividad lítica cuyo precursor
(plasminógeno) se localiza en el útero. Algunos factores expresados por el
embrión al llegar a la cavidad uterina promueven su conversión en la forma
activa.
PERIODO
IMPLANTATORIO
Consiste en una unión mucho más fuerte entre el trofoectodermo
del blastocito y las células deciduales. En esta fase se ven implicadas
numerosas moléculas de diferente naturaleza de las que cabe destacar:
·
L-selectinas: proteínas
de unión a oligosacáridos expresadas por todo la superficie del blastocito,
interacciona con los azúcares expresados en las células endometriales y permite
que el blastocito se vaya desplazando por la superficie uterina.
·
Mucina asociada a membrana (MUC-1): es
una glicoproteína de gran peso molecular que forma parte de la matriz
epitelial. Existe controversia respecto a su papel en la implantación ya que
una corriente mantiene que dificultan la adhesión debido a su gran volumen,
otra corriente piensa que puede ser un posible sustrato de las L-selectinas
expresadas en el trofoectodermo facilitando así la adhesión.
·
Integrinas: son
una amplia familia de glicoproteínas transmembrana que se unen a aquellos
ligandos que presenten la secuencia (arginina-glicina-aspártico) presente en
componentes de la matriz extracelular como la fibronectina, vitronectina, colágeno.
·
Durante la ventana de
implantación se han identificado las subunidades α1, αv, α4 y β3, apareciendo
β3 el día 5 tras la ovulación (inicio ventana implantación) y desapareciendo α4
el día 10 postovulatorio (final ventana implantación). Además el dímero αv β3
debe ser importante para la implantación, ya que su escasez dificulta este
proceso.
·
También hay una expresión
de diferentes tipos de integrinas como la β3 en el trofoblasto del embrión. Son
las encargadas de llevar a cabo una unión fuerte.
·
Fibronectina y laminina: proteínas
de la matriz extracelular que se expresan en el endometrio promovidas por la
progesterona. La laminina facilita la invasión del trofoblasto. la fibronectina
media la migración y adhesión a las células de la decidua e inhibe la
invasividad del trofoblasto.
·
Factor de crecimiento epidérmico de
unión a heparina (HB-EGF): en el endometrio humano se
expresa ampliamente en la ventana de implantación mediado por la acción tanto
de los estrógenos como de la progesterona y parece favorecer la implantación
jugando un papel importante en la iniciación de la implantación (aún es
estudio).

Implantación
Aproximadamente dos
días después de llegar el blastocisto al útero, entre seis y siete días después
de la fecundación, el blastocisto se aposiciona y adhiere al epitelio endometrial del útero, por lo general en el
polo más cercano a la masa celular interna. Tan pronto se fija al útero, el
trofoblasto comienza rápidamente a proliferar formando, gradualmente, dos
capas: el citotrofoblasto hacia adentro y el sincitiotrofoblasto del lado externo.

Disco
bilaminar
En el comienzo de la segunda semana de desarrollo
embrionario, tras el contacto del blastocisto con el endometrio se produce la
proliferación del trofoblasto del polo embrionario dando lugar a una masa de
células sin membrana conocidas con el nombre de sincitiotrofoblasto. Por el
contrario, las células del trofoblasto que forman la pared del blastocisto
conservan sus membranas constituyendo el citotrofoblasto. Cabe destacar la
actividad enzimática del sincitiotrofoblasto que degrada la matriz existente en
las células endometriales conforme aumenta el tamaño del embrión.

A medida
que la implantación del embrión progresa, el sincitiotrofoblasto rodea
gradualmente al blastocisto, hasta que finalmente, en el noveno día, todo el
blastocisto queda recubierto excepto el pequeño agujero por el cual se implantó
en la pared endometrial, que en último lugar queda sellado por material
acelular y que constituye el tapón
Alrededor
del octavo día, el embrioblasto se diferencia en dos capas; una capa externa de
células cilíndricas denominada epiblasto o ectodermo primarioy una capa interna
de células cúbicas llamada hipoblasto o endodermo prim ario. Al embrioblasto
bilaminar se le denomina disco embrionario bilaminar. Asimismo, en el mismo día
comienza a acumularse líquido entre las células del epiblasto desplazando a un
grupo de células ectodérmicas hacia el polo embrionario y constituyendo una
fina membrana denominada membrana amniótica.
La nueva
cavidad que dicha membrana delimita recibe además el nombre decavidad amniótica.
Posteriormente,
del hipoblasto emigra una capa de células que recubren la parte interna del
citotrofoblasto y que forman una membrana denominada membrana de Heuser o
exocelómica. Es por tanto cuando la cavidad del blastocisto o blastocele recibe
ahora el nombre de saco vitelino primitivo o cavidad exocelómica. Al mismo
tiempo que se forma el saco vitelino primitivo, se secreta una capa de material
acelular entre la membrana de Heuser y el citotrofoblasto denominada retículo
extraembrionario. Hacia el día 12 o 13 proliferan células procedentes del
epiblasto del extremo caudal del disco embrionario y emigran para formas dos
capas: una que recubre la superficie externa de la membrana de Heuser y otra
que recubre la superficie interna del citotrofoblasto, de modo que el retículo
extraembrionario queda atrapado entre estas dos capas donde posteriormente se
degradará y pasará a constituir la cavidad coriónica.
De nuevo,
alrededor del día 12, las células del hipoblasto comienzan de nuevo a
proliferar emigrando hacia fuera y empujando al saco vitelino primitivo hacia
el polo extraembrionario. Es entonces cuando el saco vitelino primitivo se
desprende del embrión y se desintegra formando los quistes exocelómicos, que
finalmente degenerarán. El nuevo espacio que se origina recibe el nombre de
saco vitelino definitivo o secundario. Alrededor del noveno día, las
denominadas lagunas trofoblásticas se abren en el interior del
sincitiotrofoblasto permitiendo que las sangres materna y fetal entren en
contacto al fluir por la placenta. Más adelante, los capilares maternos
próximos al sincitiotrofoblasto se expanden para formar los sinusoides
maternosque se fusionan rápidamente con las lagunas trofoblásticas. Finalmente,
el mesodermo extraembrionario induce al citotrofoblasto en su crecimiento hacia
el interior del sincitiotrofoblasto dando como resultado unas proyecciones
denominadas vellosidades primitivas.
Hacia el día 16, el mesodermo extraembrionario
asociado al citotrofoblasto penetra en las vellosidades primitivas
transformándolas en vellosidades troncales secundarias. Será más adelante, al
final de la tercera semana, cuando el mesodermo vellositario se diferencie en
los vasos sanguíneos que conecten con los vasos del embrión estableciendo una
circulación uteroplacentaria. Las vellosidades que contienen los vasos
sanguíneos diferenciados reciben el nombre de vellosidades terciarias.
Gastrulación y Formación del Disco Trilaminar
Hacia el
decimosexto día del desarrollo embrionario, cerca del centro del disco
embrionario se forma una depresión profunda rodeada por un reborde de células
epiblásticas. Este surco recibe el nombre de surco primitivo; la depresión es
la fosita primitiva y el reborde que la rodea se denomina nódulo primitivo.
Este conjunto de estructuras reciben el nombre de línea primitiva.
En el
extremo del disco embrionario, cerca de la superficie del epiblasto, se formará
la cabeza del futuro embrión. Son las células del epiblasto próximas a la línea
primitiva las que comienzan a proliferar y finalmente penetran en el espacio
existente entre el epiblasto y el hipoblasto. Este proceso se denomina gastrulación.

En él,
algunas de las células epiblásticas que penetran por la línea primitiva invaden
el hipoblasto sustituyéndolo por una nueva capa de células: elendodermo.
Asimismo, posteriormente las mismas células epiblásticas divergen en el espacio
existente entre el epiblasto y el endodermo formando una tercera capa: el
mesodermo intraembrionario.
Las células que emigran a través de la fosita
primitiva y quedan en reposo en la línea media forman dos estructuras: una masa
compacta de mesodermo craneal denominada placa precordal y un denso tubo
situado en la línea media llamado proceso notocordal. Una vez completado el
proceso de gastrulación, el epiblasto recibe el nombre de ectodermo. Por tanto,
el nuevo disco embrionario trilaminar deriva en su totalidad del epiblasto. En
el mismo día, la línea primitiva regresa en sentido caudal haciéndose cada vez
más corta hasta que finalmente desaparecerá alrededor del vigesimosexto día.
Asimismo, hacia el día 20 la línea produce una masa de mesodermo en la porción
caudal del embrión denominada eminencia caudal, a partir de la cual surgen
estructuras mesodérmicas caudales del organismo.
Cuando el
proceso notocordal se ha formado por completo, lo que sucede alrededor del día
20, ocurren los siguientes cambios estructurales: - El suelo ventral del tubo
se fusiona con el endodermo subyacente. - El tubo se abre ventralmente
comenzando por la región de la fosita primitiva. Todos estos cambios convierten
al proceso notocordal en una barra medio ventral aplanada de mesodermo
denominada placa notocordal. Dicha placa cambiará de forma poco a poco y
acabará convirtiéndose en dos o tres días en un cilindro sólido denominado
notocorda. En este proceso, algunas células endodérmicas pueden quedar
incorporadas a la notocorda. La función que desempeña la notocorda no está
claramente definida pero se conoce que desempeña un papel importante en la
inducción de los cuerpos vertebrales.
De modo
transitorio, la cavidad del saco vitelino comunicará con la cavidad amniótica a
través de una apertura en la fosita primitiva denominada conducto
neuroentérico. Durante esta tercera semana, aparecen dos depresiones en el
ectodermo: una en el extremo craneal denominada membrana bucofaríngea y otra en
el extremo caudal denominada membrana cloacal. Ambas membranas de origen
ectodérmico comunicarán con el endodermo subyacente excluyendo al mesodermo
intra embrionario y formando por tanto una membrana bilaminar. Ambas membranas
se convertirán además en los extremos ciegos del tubo digestivo.
Conforme
la línea primitiva regresa en sentido caudal, las células mesodérmicas que
emigran lateralmente a partir de ella, comienzan a condensarse en estructuras
laminares o cilíndricas a ambos lados de la notocorda. Este proceso se inicia
en el extremo craneal y avanza en sentido caudal. El mesodermo situado
inmediatamente a ambos lados de la notocorda forma un par de condensaciones
cilíndricas denominadas mesodermo paraxial. Asimismo, un par de condensaciones
menos pronunciadas aparece junto al mesodermo paraxial denominadas mesodermo
intermedio. Finalmente, el resto de mesodermo lateral compone una lámina que
recibe el nombre de mesodermo lateral. A partir del día 17, el mesodermo
lateral se divide en dos capas: una capa adyacente al endodermo que recibe el
nombre de mesodermo esplacnopleural y una capa adyacente al ectodermo denominada
mesodermo somatopleural.
Al mismo tiempo, tan pronto como
se forma el mesodermo paraxial, éste acaba produciendo un conjunto de
estructuras redondeadas denominadas somitámeros. Dichas estructuras derivan del
mesodermo paraxial y avanzan formándose en sentido cráneo caudal. La mayoría de
los somitámeros se diferencian más tarde para formar bloques separados
denominados somitas. Sin embargo, cabe destacar que los siete primeros pares de
somitámeros no desarrollan somitas sino que suelen dar lugar a otras
estructuras craneales como la mandíbula, los músculos de la cara, etc.
Finalmente, el recuento de somitas en el ser humano suele rondar el número de
37 pares.
En la segunda semana se
ve el desarrollo de órganos importantes en el embrión y las posibles mal
deformaciones que se pueden presentar en esta semana, el sistema nervioso
comienza su desarrollo embriológico, se da forma a la línea media de la región
dorsal del embrión la membrana bucofaríngea y el nodo primitivo.
TEJIDOS
ECTODERMO
El ectodermo es la capa más externa (distal). Es la primera en formarse,
durante la fase de blástula del
desarrollo embrionario y más adelante da lugar a las otras dos durante la gastrulación.
el ectodermo se diferencia para formar el sistema nervioso
(médula espinal, nervios periféricos y cerebro), el esmalte dental
y la epidermis (las
partes externas del integumento). También forma el revestimiento de la boca, ano, fosas
nasales, glándulas sudoríparas, pelo y uñas.
Emerge primero del epiblasto
durante la gastrulación y forma la capa externa de las capas germinativas.

Es el que actúa de
superficie del cuerpo, en superficies externas como cavidades y conductos
internos. Como: la piel, mocosas y glándulas.
En algunos órganos, lo
encontramos como el parénquima y en cavidades internas como el endotelio.
Clasificación epitelial por su origen
-
Epitelio ciliado
Los cilios son
proyecciones largas de la membrana plasmática hacia la luz del conducto o
cavidad que recubre el epitelio.
Cada célula tiene
muchos cilios que cuentan con movilidad ya que ejercen su acción de transporte.
Mueven el moco o
líquido a traves de conductos y órganos tubulares.Se mueven de forma coordinada
y rítmica.
-
Epitelio flagelado
Función de transporte y
función sensorial en los epitelios sensoriales.
Número menor de cilios
por célula y el movimiento es más independiente del resto.
-
Epitelio con microvellosidades
Las microvellosidades
son especializadas de la membrana plasmática apical en epitelios de absorción.
Su función es aumentar
la superficie de absorción.
Se encuentra en algunas
partes del riñon y aparato digestivo.
Los esterocilios son
estructuras asociadas a las
microvellosidades que no tienen movilidad.

Clasificación epitelial por su
estructura
Estructura simple
•
1. EPITELIO SIMPLE PLANO: está compuesto por una capa
única de células planas firmemente unidas. Las células presentan un núcleo
prominente y aplanado, Se encuentra en los vasos sanguíneos y linfáticos, en la
cubierta del ovario, en los alvéolos pulmonares, el asa de Henle, la cápsula de
Bowman y también el mesotelio de las serosas. Función principal es de
intercambio y lubricación.
•
2.
EPITELIO SIMPLE CÚBICO: Posee 2 capas de células cúbicas. Funcion de
absorción y secreción. La capa de células unidas de forma cúbica con un núcleo
redondo ubicado en el centro, reviste los ductos de muchas glándulas endocrinas
(tiroides, por ejemplo), y ductos del riñón (túbulos renales) y la capa
germinativa de la superficie del ovario.
•
3. EPITELIO SIMPLE CILÍNDRICO: Función de absorción y la
secreción, constituye el revestimiento del intestino desde el cardias hasta el
ano, también se encuentra en la vesícula biliar y los conductos mayores de las
glándulas. Presentan un núcleo ovoide. Pueden presentar un borde estriado o
microvellosidades. Reviste el útero, oviductos, conductos deferentes, pequeños
bronquiolos y senos paranasales es ciliado.
Epitelio estratificado o
poliestratificado
•
1. EPITELIO ESTRATIFICADO PLANO Las células de la capa superficial son planas y
pueden ser o no queratinizados. El epitelio plano estratificado queratinizado
es el que forma la capa externa de la piel, la epidermis. La última capa está
formada por células muertas con el interior celular lleno de queratina. El
epitelio plano estratificado no queratinizado se puede encontrar en mucosas,
como la lengua, esófago, faringe o vagina.
•
2. EPITELIO ESTRATIFICADO COLUMNAR: Este es uno de los tipos de epitelio poco
frecuentes. Suele tener una función protectora y se puede encontrar en algunas
zonas de la faringe, en algunas zonas de la uretra masculina y otros conductos excretores
mayores de ambos sexos y en la conjuntiva del ojo.
•
3. EPITELIO ESTRATIFICADO cúbico Las glándulas sudoríparas son el único sitio del
cuerpo humano dónde aparece este tipo de epitelio que está formado por dos
capas de células cúbicas siendo las inferiores de mayor volumen.
Epitelio pseudoestratificado
•
Se caracteriza
por contar con una sola capa de células si se mira desde la membrana basal pero
que puede dar la sensación de múltiples capas si se observa desde la
superficie. Esto se debe a que no todas las células alcanzan la superficie
pero todas alcanzan la membrana basal su función es la secreción y absorción.
Epitelio glandular:
•
GLÁNDULAS EXOCRINAS
•
En las glándulas exocrinas, el producto
celular va a llegar a la superficie a través de los conductos.
•
Su unidad celular es el Adenomero
•
El estroma es el tejido conectivo que
sostiene y recubre a la glándula
•
GLÁNDULAS ENDOCRINAS
•
No tienen conducto externo
•
Vierten la secreción directamente en la
linfa y en la sangre

MESODERMO
El mesodermo es
una de las tres capas celulares embrionarias que surgen durante el proceso de
gastrulación, alrededor de la tercera semana de gestación.
Desarrollo del mesodermo y sus
derivados
El mesodermo da lugar
principalmente a los músculos, huesos y vasos sanguíneos. Durante las primeras
etapas del desarrollo embrionario, las células forman dos clases de tejidos:
Epitelios:
las células se conectan a través de fuertes uniones construyendo láminas. El
mesodermo forma numerosos epitelios.
Mesénquima:
las células se distribuyen dejando amplios espacios entre ellas, constituyendo
un tejido de relleno. La mesénquima es el tejido conectivo, y gran parte de
ella proviene del mesodermo. Una pequeña
parte surge del ectodermo.

-
Éste corresponde a una estructura
fundamental en el desarrollo llamada notocorda. Ésta tiene forma de cordón, y
se localiza en la línea media de la parte dorsal del embrión. Es el eje de
referencia que determinará que ambos lados del cuerpo se desarrollen de forma
simétrica.
-
La notocorda empieza a formarse a los 18
días de gestación, mediante unos movimientos celulares ocurridos durante el
periodo de gastrulación. Comienza con una grieta superficial que se va
plegando, y se invagina hasta formar un cilindro alargado.
Mesodermo paraxial
-
Es la parte
más gruesa y ensanchada del mesodermo. Sobre la tercera semana, se divide en
segmentos (llamados somitámeros) que van apareciendo en orden cefálico a
caudal.
Mesodermo lateral
-
Es la parte
más externa del mesodermo. A los 17 días de gestación aproximadamente, el
mesodermo lateral se divide en dos láminas: el mesodermo esplacnopleural, que
está al lado del endodermo; y el mesodermo somatopleural, que se localiza
adyacente al ectodermo.
TEJIDO MUSCULAR
Es un tejido formado por células de
la misma especie, estas células son llamadas células contráctiles (miocitos)
que se han especializado al máximo para conseguir un correcto funcionamiento
mecánico a partir de la energía química gracias a la interacción de las
proteínas contráctiles (actina y miosina).
El músculo liso, también conocido como no curvo o
no voluntario, se compone de células en forma de huso.
El estímulo para la contracción de los músculos
lisos está mediado por el sistema nervioso autónomo.

Funciones
Facilitarla incorporación
diversos elementos dentro del cuerpo:
›
Sangre para el músculo liso de los vasos sanguíneos,
›
Aire para el músculo liso bronquial,
›
Alimento para el músculo liso del tracto digestivo
›
Orina para el músculo liso de los riñones, la vejiga
y los haces que transportan la orina.
›
De la dermis de la piel
Características
ü Forma las paredes de los
órganos
ü Involuntario
ü Tono muscular
ü Se contrae bajo diferentes
estímulos
Tipos de musculo liso
Multiunitario : Formado por fibras musculares lisas separadas y
discretas.
Cada una de las fibras actúa independientemente
de las demás.
La superficie externa esta cubierta por una capa
de sustancias similares a una membrana
basal , colágeno y glucoproteínas.
Unitario:
También
llamado musculo liso visceral.
Es una masa de cientos de miles de fibras de
musculo liso que se contraen como una
unidad única.
Las fibras están dispuestas en laminas o
fascículos.
Morfología
ü No presenta estrías
ü No tiene líneas Z
ü Pocas mitocondrias
ü No presenta troponina
ü Presenta tropomiosina
Se localiza en órganos como:
Aparato respiratorio, aparato digestivo, aparato urinario, vasos sanguíneos.
Aparato respiratorio, aparato digestivo, aparato urinario, vasos sanguíneos.
Presente en todos los
músculos de las vísceras y en todos los paneles vasculares.
Tejido Muscular Estriado
Origen embrionario
•
Se desarrolla en el tejido conectivo a partir del
mesénquima
•
Aparece en la quinta semana de la vida fetal
•
Las células mesenquimatosas indiferenciadas se
transforman en mioblastos , los cuales son de forma alargada y poseen un solo
núcleo
•
En los mioblastos se colocan uno a continuación de
otros desapareciendo las zonas del sarcolema, lo cual provoca la fusión del
citoplasma
•
Como resultado da las fibras musculares estriadas
Este tejido constituye todos los llamados
músculos esqueléticos, que quiere decir que están unidos a los huesos .
Su función es el movimiento y la producción de calor del músculo
Unidad fundamental
Fibras musculares o también llamadas miocitos.
Son cilíndricas largas y alargadas.
Sus núcleos son periféricos y se
encuentran rodeadas por una membrana denominada sarcolema.
Estructura
Fascículos.-
son unidades funcionales de fibras musculares que
actúan en conjunto para desempeñar una función específica.
Endomisio.- delicada capa de fibras reticulares que rodea las
fibras musculares individuales.
Perimisio.- Capa de tejido conjuntivo que rodea un grupo de
fibras para formar un haz o fascículo.
Epimisio.- Es la vaina de tejido conjuntivo denso que rodea
todo el conjunto de fascículos que forman el músculo.

TEJIDO
MUSCULAR CARDIACO
El tejido muscular estriado cardíaco
es un tipo especial de músculo que forma exclusivamente el
corazón. El miocardio, juntamente con el endocardio y en pericardio,
son los tres tejidos que forman el órgano. Este músculo debe ser capaz de contraerse y relajarse de forma
ininterrumpida desde antes de que nazca el animal hasta el omento de su muerte,
por lo que requiere unas fibras
que no se fatiguen con los trabajos prolongados y ha de ser capaz de hacer
mover toda la sangre por el cuerpo del individuo.
Fisiológicamente el
miocardio se caracteriza por poder transmitir el impulso nervioso, como si fuera una neurona.
Además el corazón es capaz de generar
su propio potencial eléctrico que es el responsable de su propia
contracción, al contrario que los músculos esqueléticos.
Este tejido está compuesto por cardiomiocitos. Estas células son mononucleadas, con el núcleo en
posición central y de apariencia ovoide con la cromatina poco concentrada. Al
contrario que las fibras musculares, los cardiomiocitos no tienen una forma de
huso tan pronunciada, sino que son más redondeados
y presentan ramificaciones. En su citoplasma contienen estrías de miosina y actina en una ordenación reticular similar
a la que se puede observar en las fibras musculares.

estrías de miosina y
actina en una ordenación reticular similar a la que se puede observar en las fibras
musculares.
El tejido muscular del corazón tiene dos
características histológicas que lo diferencian del estriado esquelético: El espacio perinuclear se encuentra libre de las
estriaciones de fibras de miosina y actina. Alrededor del núcleo estas
células almacenan glucógeno, que
son capaces de convertir en glucosa, como fuente de energía en caso de que no
obtengan la suficiente por la sangre. Aun así el corazón necesita un aporte constante de oxígeno para su
funcionamiento (mediante la fosforilación oxidativa), sin dicho aporte
sufre daños celulares rápidamente.
Las células del miocardio están muy estrechamente relacionadas entre sí.
Se encuentran conectadas por discos
intercalares proteicos especiales que permiten la transmisión del
impulso nervioso entre ellas. Estas uniones, con forma de hendidura, permiten la sincronización del órgano para
realizar la contracción adecuadamente.
La contracción
cardíaca se encuentra controlada, de forma involuntaria, por el 0 nervioso autónomo,
éste se encarga de controlar la fuerza de las contracciones y su frecuencia.
Aunque la contracción en sí se genera a partir de unos miocitos especializados,
el nódulo sinoauricular situados
en la pared superior de la aurícula derecha. Si bien no todo todas las células
cardiacas se encuentran inervadas con una neurona. Además las hormonas que llegan por el torrente
sanguíneo también pueden controlar el ritmo cardíaco.
ENDODERMO
El endodermo
es una de las tres capas germinales que surgen en el desarrollo embrionario
temprano, sobre la tercera semana de gestación. Las otras dos capas se conocen
como ectodermo o capa externa y mesodermo o capa media.
Una de
las capas más antiguas de la diferenciación embrionaria en los seres vivos es
el endodermo. Por ese motivo, provienen de ella los órganos más importantes
para la supervivencia del individuo.

Desarrollo del endodermo
La
diferenciación del cuerpo del embrión del líquido exterior afecta al endodermo,
dividiéndolo en dos partes: el endodermo embrionario y el extraembrionario.
Endodermo embrionario
La
transformación del endodermo va paralela a los cambios inducidos por la
notocorda. Así, la notocorda induce pliegues que van a determinar los ejes
craneal, caudal y lateral del embrión. El endodermo también se pliega
progresivamente hacia el interior de la cavidad corporal por la influencia de
la notocorda.
Endodermo extraembrionario
La otra
porción de endodermo queda fuera del embrión, y se denomina saco
vitelino. El saco vitelino consiste en una estructura
membranosa adherida al embrión que se encarga de nutrir, dar oxígeno a éste y
eliminar desechos.
Partes del tubo intestinal del
endodermo
Por otro
lado, se pueden diferenciar distintas áreas en el tubo intestinal del
endodermo. Cabe decir que algunas de ellas pertenecen al endodermo embrionario
y otras al extraembrionario:
– El
intestino craneal o interior, que se encuentra dentro del pliegue de la cabeza
del embrión. Comienza en la membrana bucofaríngea, y esta región va a convertirse
en la faringe. Luego, en el extremo inferior de la faringe aparece una
estructura que originará el tracto respiratorio.
Debajo de
esta área, el tubo se ensanchará rápidamente para convertirse posteriormente en
el estómago.
–
Intestino medio, situado entre el intestino craneal y caudal. Éste se prolonga
hasta el saco vitelino mediante el cordón umbilical. Esto permite al embrión
recibir nutrientes del organismo de su madre.
– El
intestino caudal, dentro del pliegue caudal. A partir de él surge el alantoides, una membrana
extraembrionaria que aparece por una invaginación localizada al lado del saco
vitelino.
Consiste
en un depósito que sale del cuerpo embrionario a través del pedículo del
alantoides (cordón umbilical). El volumen del líquido de la bolsa va a medida
que avanza la gestación, ya que parece ser que este saco acumula los desechos
metabólicos.
En los
seres humanos, el alantoides da lugar a los vasos umbilicales y a las vellosidades
de la placenta.
Derivados del endodermo
Como se
mencionó, el endodermo deriva en órganos y estructuras del cuerpo a través de
un proceso llamado organogénesis. La organogénesis ocurre en una etapa que dura
desde la tercera a la octava semana de gestación aproximadamente.
El
endodermo contribuye en la formación de las siguientes estructuras:
–
Glándulas del tracto gastrointestinal y órganos gastrointestinales asociados
como el hígado, la vesícula biliar y el páncreas.
–
Epitelio o tejido conectivo que rodea: las amígdalas, la faringe, la laringe,
la tráquea, los pulmones, y el tracto gastrointestinal (menos la boca, el ano,
y parte de la faringe y el recto; que provienen del ectodermo).
También
forma el epitelio de la trompa de Eustaquio y la cavidad timpánica
(en el oído), las glándulas tiroideas y paratiroideas, la glándula timo, la
vagina y la uretra.
– Tracto
respiratorio: como bronquios y alvéolos pulmonares.
– Vejiga urinaria.
–
Saco vitelino.
–
Alantoides.
Se ha
visto que en los seres humanos el endodermo puede diferenciarse en órganos
observables tras las 5 semanas de gestación.
Tejido
Conectivo
Es un conjunto heterogéneo de tejidos orgánicos que comparten un origen común a
partir del mesénquima embrionario originado a partir del mesodermo.
Su
función de relleno, ocupando los espacios entre otros tejidos y entre órganos,
y de sostén del organismo, constituyendo el soporte material del cuerpo.

Su
estructura. Están formados por:
- Células bastante separadas entre sí. Se denominan con la terminación “-blasto” cuando tienen capacidad de división y fabrican la matriz intercelular y con la terminación “-cito” cuando pierden la capacidad de división.
- Fibras de colágeno (proporcionan resistencia a la tracción), de elastina (proporcionan elasticidad) y de reticulina (proporcionan unión a las demás estructuras).
- Matriz intercelular de consistencia variable que rellena los espacios entre células y fibras y constituida por agua, sales minerales, polipéptidos y azúcares. La consistencia de la matriz determina la clasificación de los tejidos conectivos.
- Células bastante separadas entre sí. Se denominan con la terminación “-blasto” cuando tienen capacidad de división y fabrican la matriz intercelular y con la terminación “-cito” cuando pierden la capacidad de división.
- Fibras de colágeno (proporcionan resistencia a la tracción), de elastina (proporcionan elasticidad) y de reticulina (proporcionan unión a las demás estructuras).
- Matriz intercelular de consistencia variable que rellena los espacios entre células y fibras y constituida por agua, sales minerales, polipéptidos y azúcares. La consistencia de la matriz determina la clasificación de los tejidos conectivos.
Tejido
Conectivo Denso
•
Estructura
•
Tipo de
tejido conectivo, que se encuentra dentro de los tejidos conjuntivos .
•
Forma
los tendones, aponeurosis, ligamentos y en general estructuras que reciben
tracción en la dirección hacia la cual se orientan sus fibras colágenas.
•
Estas
fibras se hallan dispuestas en una forma ordenada, paralela una respecto a la
otra, lo que proporciona la máxima fortaleza.
•
Función
•
Rellena
los espacios comprendidos entre los diferentes tejidos y estructuras.
•
Reserva
energética en las células adiposas.
•
Participa
en la defensa del organismo a través de sus células.
•
Produce
células sanguíneas en la médula ósea.
•
Clasificación
•
Conjuntivo
denso irregular: Se localiza en la capa inferior (dermis) de la piel. Las
fibras colágenas son más gruesos y están dispuestos irregularmente y entre
tejidos. Las fibras colágenas están asociadas con redes de fibras elásticas.
•
Conjuntivo
denso regular: Se localiza en los ligamentos, tendones y aponeurosis.
Macroscópicamente este tejido muestra una estructura perceptiblemente fibrosa y
un aspecto característico, debido a su color blanco brillante. Los únicos
elementos celulares presentes son los fibroblastos, los cuales se disponen
entre los haces paralelos de las fibras colágenas.

Tejido
Cartilaginoso
•
El tejido
cartilaginoso (o cartílago) pertenece a la categoría de los tejidos conjuntivos
de soporte. Las células del tejido cartilaginoso son denominadas condrocitos,
que están localizados en lagunas (o condroplastos) de la matriz.
•
Las células
que lo componen, los condroblastos, forman la matriz extracelular. Sus
células maduras, los condrocitos, forman grupos aislados unos de otros en
unas cavidades llamadas lagunas.
•
La nutrición
del tejido cartilaginoso está asegurada por los capilares del tejido conjuntivo
envolvente, puesto que no posee capilares sanguíneos.
•
Este tipo de
tejido tampoco contiene vasos linfáticos ni nervios.

Funciones
•
Soporte de los
tejidos blandos
•
Recubrimiento
de superficies articulares
•
Facilidad del
deslizamiento de los huesos en las articulaciones
•
Fundamental
para la formación y crecimiento de los huesos largos durante el desarrollo
fetal y tras el nacimiento.
Clasificación
•
Cartílago
hialino
•
está envuelto
por una capa de tejido conjuntivo, el pericondrio, que puede dar origen a
nuevos condrocitos.
•
Se encuentra
en la tráquea, en los bronquios, en la extremidad ventral de las costillas y en
las superficies articulares de los huesos largos.
•
Cartílago
elástico
•
Abundancia de
fibras elásticas, además de poseer fibrilas de colágeno (principalmente del
tipo II). También está envuelto por el pericondrio, siendo el principal
responsable por el crecimiento de este tipo de cartílago.
•
Se encuentra
en el pabellón auditivo, en el canal auditivo externo, en la laringe y en la
trompa de Eustaquio.
•
Cartílago
fibroso
•
Abundancia de
fibrilas colágenas (del tipo I) . Tiene poca sustancia fundamental, sin
pericondrio, asociada al tejido conjuntivo denso
•
Se encuentra
en los discos intervertebrales y el cartílago de los meniscos.
Tejido
conectivo laxo
El tejido conectivo laxo es un tipo de tejido muy abundante en el
organismo, cuyo origen proviene del mesenquima. Las células del mesodermo son
pluripotenciales, dando lugar a otros tipos celulares, como son el tejido conjuntivo, tejido cartilaginoso, tejido óseo y tejido
cordal. Contienen un alto porcentaje
de fibras (predomina la fibra de
colágeno),
algunos fibroblastos, macrófagas y sustancia fundamental de la
matriz extracelular.

CLASIFICACION
Las fibras de tejido laxo se encuentran entrelazadas en el espacio
intracelular. Existen tres tipos de tejido laxo:
- Tejido
conectivo areolar: Es de los de más amplia
distribución. Contiene tipos celulares como fibroblastos, macrófagos,
células plasmáticas, mastocitos y glóbulos blancos. Se compone de fibras
de colágena, elásticas y reticulares. Las sustancias principales que
contiene son ácido hialurónico, condroitin sulfato. Este tipo de tejido
forma parte del tejido subcutáneo.
- Tejido
adiposo: Se compone de un tipo célular llamado adipocira que almacenan
triglicéridos. Se encuentra asociado al tejido areolar. Actúa como
aislante para evitar la pérdida de calor. Es la principal fuente de
energía y brinda soporte y protección a diversos órganos.
- Tejido
conectivo reticular: Es un conjunto de
células y fibras reticulares que constituyen el estroma de algunos órganos
como el hígado, bazo y ganglios linfáticos. Las células de este tejido
filtran la sangre eliminando las células viejas
Localización
El tejido conectivo laxo está muy vascularizado. Este se encuentra
localizado debajo de los epitelios, en la mucosa y submucosa de la pared del
tubo digestivo, del sistema urinario y del respiratorio.
Función
El tejido conectivo laxo actúa como un soporte y alineador celular y
hormonal ya que eyacula una sustancia llamada segregina, encargada de la
reproducción de hormonas. Forma la dermis de la piel, rellena y rodea numerosos
órganos, además de nutrirlos. También poseen una función defensiva.
Tejido adiposo
El tejido adiposo o tejido graso es el tejido de origen mesenquimal (un tipo de tejido conjuntivo) conformado por la asociación
de células que acumulan lípidos en su citoplasma: los adipocitos.
El tejido adiposo, por un lado, cumple funciones mecánicas: una de ellas
es servir como amortiguador, también protegiendo y manteniendo en su lugar a
los órganos internos así como a otras estructuras externas del cuerpo, y
también tiene funciones metabólicas y es el encargado de generar grasas para el
organismo.

Existen dos tipos de tejido adiposo, el tejido
adiposo blanco (o unilocular) y la grasa parda (o multilocular).
El citosol y el núcleo quedan reducidos a una pequeña área cerca de la membrana. El resto es ocupado por una
gran gota de grasa. El tejido adiposo, que carece de sustancia fundamental, se
halla dividido por finas trabéculas de tejido
fascicular en
lóbulos.
La grasa de las células se encuentra en estado semilíquido y también
está compuesta fundamentalmente por triglicéridos. Se acumula de preferencia en
el tejido subcutáneo, la capa más profunda de la piel. Sus células, lipocitos, están especializadas en formar
y almacenar grasa. Esta capa se denomina panículo adiposo y es un aislante del frío y del calor. Actúa como una almohadilla y
también como un almacén de reservas nutritivas.
Este tipo de tejido cumple funciones de relleno y de amortiguación,
especialmente en las áreas subcutáneas. También sirve de soporte estructural y
una función de reserva energética. La grasa varía de consistencia, es decir
puede ser encontrada tanto en estado líquido como sólido.
El crecimiento de este tejido se puede producir por proliferación
celular (crecimiento hiperplásico), por acumulación de una mayor cantidad de lípidos en las células ya existentes
(crecimiento hipertrófico) pero nunca aumenta el número de adipocitos por
división mitótica. Durante la adolescencia el crecimiento es,
generalmente, rápido y en el individuo adulto hipertrófico.
Características anatómicas
En los humanos, el tejido adiposo está
localizado debajo de la piel (grasa subcutánea), alrededor
de los órganos internos (grasa visceral), en la médula ósea (médula ósea amarilla) y en
las mamas. El tejido adiposo está localizado en regiones específicas, las
cuales se conocen como depósitos de adipocitos. Además de los adipocitos, que conforman el
porcentaje más alto de células en el tejido adiposo, existen otras células que
están presentes de manera colectiva denominadas fracción de estroma visceral
(SVF). Este estroma está formado por pre-adipocitos, fibroblastos, macrófagos
de tejido adiposo, y células endoteliales. El tejido adiposo contiene pequeños
vasos sanguíneos.
En el sistema tegumentario, el cual incluye la piel, el tejido adiposo
se almacena en la capa más profunda de la piel regulando la temperatura del
cuerpo.
Alrededor de los órganos, éste tejido brinda protección. Sin embargo su
función principal es ser una reserva de lípidos, los cuales pueden ser
utilizados para generar la energía necesaria para el cuerpo y protegernos del
exceso de glucosa.1 Bajo condiciones normales, brinda estímulo de hambre y saciedad al
cerebro.
Fisiología
Los ácidos grasos libres son liberados de la lipoproteína por una enzima
llamada lipasa lipoproteíca; éstos ácidos grasos libres entran al adipocito,
donde son reensamblados en triglicéridos. Aproximadamente el 87% del tejido
graso de los humanos está compuesto por lípidos.
Existe un constante flujo de ácidos grasos libres. Dichos fluidos son
controlados por la insulina y la leptina. Si tenemos una concentración elevada
de insulina existe un incremento en el flujo de ácidos grasos libres, cuando la
insulina baja, los ácidos grasos pueden ser liberados del tejido adiposo. La
secreción de insulina es estimulada por la concentración elevada de azúcar o
glucosa en sangre debido al consumo de carbohidratos.
En humanos, la lipólisis (hidrólisis de triglicéridos en ácidos grasos)
es regulada por el balance controlado de los receptores andrógeno-B lipolítico
y el receptor androgénico a2A, mediando la anti-lipólisis. Los adipocitos
tienen un papel fisiológico importante en la regulación de los niveles de los
triglicéridos y los ácidos grasos libres, así mismo determinan la resistencia a
la insulina.
La grasa abdominal tiene un metabolismo diferente, siendo más propenso a
inducir la resistencia a la insulina. Esto explica porque la obesidad central
es un precursor de la intolerancia a la glucosa siendo un factor independiente
a enfermedades cardiovasculares (aún en la ausencia de diabetes mellitus e hipertensión).
Tejido óseo
El tejido óseo es
un tejido especializado del tejido conjuntivo, constituyente principal de los huesos en los vertebrados. Está compuesto por células y componentes extracelulares calcificados que forman la matriz ósea. Se caracteriza por su rigidez y su gran
resistencia a la tracción, compresión y a las lesiones.
Estructura histológica
La estructura histológica del tejido
óseo maduro es igual tanto en la sustancia compacta como en la sustancia
esponjosa y se designa con el nombre de hueso laminar. Durante el desarrollo se
forma hueso entretejido o inmaduro que luego se transforma en hueso laminillar.
Las unidades estructurales del
tejido óseo maduro son nominillas óseas de 3 a 7 µm de espesor
(especiales o concéntricas) que sobre todo en las regiones de sustancia
compacta forman sistemas tubulares finos, las osteonas. El hueso laminillar puede estudiarse con la ayuda de
dos técnicas de preparación diferentes: Artrosis. Es cuando se gasta el líquido
sinovial.
1.
Método de desgaste: A partir del hueso macerado limpio,
en los que se puede ver sobre todo la distribución y la organización del
material inorgánico con calcio abundante.
2.
Corte de espécimen descalcificado: Muestra el material
orgánico (colágeno, células y vasos).
Sistema de
laminillas
La mayoría de las laminillas están orientadas en
forma concéntrica alrededor de los vasos que transcurren longitudinalmente y
aquí forman unidades estructurales cilíndricas, las osteonas o sistemas de
Havers. En el corte transversal las osteonas tienen forma redondeada oval y
también irregular, en ocasiones son ramificadas y pueden anastomosarse entre
sí. Su diámetro varía entre 100 y 400 µm. Pueden alcanzar algunos
centímetros de largo. La cantidad de laminillas de una osteona oscila entre los
4 y 20. Las laminillas de las osteonas también se denominan laminillas
especiales. En ellas transcurren fibrillas colágenas con una disposición
helicoidal. Entre las osteonas hay restos de osteonas antiguas y degradadas que
llenan el espacio entre las osteonas intactas como con "balasto"
(laminillas intersticiales, sistemas intersticiales). Los límites entre las
osteonas y los sistemas intersticiales están marcados con nitidez por las
denominadas líneas de cemento (contienen muchos proteoglucanos). En sus
superficies externas e interna, las regiones óseas compactas se caracterizan
por poseer laminillas que rodean todo el elemento óseo (por fuera) o tapizan la
cavidad medular (por dentro) y reciben los nombres de laminillas
circunferenciales externas e internas.
Células del hueso
Este tejido se renueva y se
reabsorbe continuamente, gracias a la actividad de sus células específicas.
Estas son los osteoblastos, responsables de la formación de tejido
óseo nuevo; los osteocitos, que son los osteoblastos maduros y
desarrollan una actividad menor; y los osteoclastos, que se
encargan de reabsorber o eliminar la materia ósea.
Células osteoprogenitoras
·
El tejido óseo se origina a partir de células de origen
mesenquimal (como todos los tejidos conectivos). A partir de las células
mesenquimales que se comprometen hacia una diferenciación en células formadoras
de hueso se forma una colonia celular con potencial más limitado para
proliferar y diferenciarse, estas son las células osteoprogenitoras.
·
Las células osteoprogenitoras cuentan con potencial para
diferenciarse hacia condroblastos u osteoblastos.
·
Las células osteoprogenitoras persisten hasta la vida
postnatal y se hallan en casi todas las superficies libres de los huesos
(endostio, capa interna del periostio, trabéculas de cartílago calcificado).
·
Durante la fase de crecimiento de los huesos, las células
osteoprogenitoras son más activas; aunque también aumenta su actividad ante el
fenómeno de reparación de lesiones óseas.
·
Como se supone que los osteoblastos y los osteocitos
carecen de capacidad mitótica, parece evidente que a medida que disminuye la
población de osteoblastos durante los procesos de remodelación continua del
hueso, las células osteoprogenitoras que proliferan y se diferencian proveen de
nuevos osteoblastos para el tejido.
Osteoblastos
Son células osteoformadoras que se
encargan del mantenimiento, el crecimiento y la reparación del hueso.
Son los encargados de sintetizar los
componentes de la matriz ósea.
Están provistos de un retículo
endoplasmático y un aparato de Golgi muy desarrollados. Además sus mitocondrias
concentran gránulos de fosfato de calcio. Estos van a ser transferidos a
vesículas que posteriormente pasarán al medio extracelular para poder acumular
fosfatos y calcio por medio activo. Se acumulan progresivamente hasta que se
rompen, y las partículas liberadas se fijan a las partículas de colágeno en
forma de cristales de hidroxihapatita, produciendo la mineralización de la
matriz.
Una vez que los osteoblastos están
rodeados por la matriz, pasarán a denominarse como osteocitos.
Osteocitos
Son las células del hueso maduro y
ya formado y se presentan bajo tres estados funcionales: osteocitos
latentes, osteocitos formativos y osteocitos
resortivos.
Se localizan en lagunas y emiten
prolongaciones a través de los conductos calcóforos (canales presentes en la
matriz). Se comunican unos con otros a través de uniones comunicantes como son
los conductos.
Estas células poseen la capacidad de
síntesis y reabsorción de la matriz.
Osteoclasto
El osteoclasto es una célula
multinucleada que degrada y reabsorbe huesos. Al igual que el osteoblasto, está
implicado en la remodelación de hueso natural.
Son células multinucleadas y
polarizadas. La zona de las células que no está en contacto con el hueso es
pobre en orgánulos y posee filamentos de actina. También es donde se encuentran
los núcleos. La otra parte de las células que está en contacto con el hueso se
caracteriza por la presencia de numerosas prolongaciones citoplasmáticas que se
denominan borde plegado o borde fruncido.
Tipos de tejido óseo

Prosaicamente se distinguen dos
zonas óseas con características diferentes y sin un límite neto, éstas
representan dos formas diferentes de estructuración del tejido óseo:
Tejido esponjoso o areolar
El hueso esponjoso constituye la
mayor parte del tejido óseo de los huesos cortos, planos y de forma irregular y
de la epífisis de los huesos largos. El tejido esponjoso de los huesos de la
pelvis, las costillas, el esternón, las vértebras, el cráneo y los extremos de
algunos huesos largos es el único reservorio de médula ósea roja y por lo
tanto, de hematopoyesis en los adultos. El hueso esponjoso no contiene
verdaderas osteonas. Está formado por laminillas dispuestas en un encaje
irregular de finas placas de hueso llamadas trabéculas. Los espacios entre las
trabéculas de algunos huesos están ocupados por la médula ósea roja productora
de células sanguíneas. En el interior de las trabarlas existen eritrocitos,
situados en lagunas de las que parten conductillos radiales. Los vasos
sanguíneos del periostio penetran a través del hueso esponjoso. Los eritrocitos
de las trabarlas reciben su nutrición directamente de la sangre que circula por
las cavidades medulares.
Tejido compacto o denso
Se encuentra en la capa externa de
los huesos largos formando la diáfisis, en el exterior y en el interior de los
huesos planos y en distintas zonas en los huesos cortos, según cada hueso en
concreto. Es un tejido duro, denso y frágil. Al observarlo al microscopio
destacan estructuras cilíndricas, denominadas osteonas, formadas por capas
concéntricas de laminillas óseas, donde se encuentran insertos los osteocitos.
En el interior de la osteona hay un canal, el conducto de Havers, por donde
circulan vasos sanguíneos, linfáticos y nervios. Los vasos sanguíneos aportan
los nutrientes necesarios a las células de los huesos y conducen las hormonas
que controlan el aporte de calcio. También aparecen canales que conectan unos
conductos de Havers con otros. Estos conductos se llaman conductos de Volkmann.
La unidad estructural del tejido óseo compacto es el Sistema de Aversar u
osteona. Este tipo de hueso se localiza en la diáfisis de los huesos largos. El
Sistema de Havers está constituido por:
·
Conducto de Aversar: contiene el VAN que pasa por el
agujero nutricio del hueso.
·
Laminillas óseas: su número aumenta a medida que crece el
hueso.
·
Osteoplastos: dispuestos en forma concéntrica, en cuyo
interior se encuentran los osteocitos.
·
Canalículos calcóforos: conecta el osteoplasto con el
conducto de Havers. Permite la nutrición y eliminación de desechos del
osteocito (sus prolongaciones viajan a través del canalículo).
ORTOBIOLOGIA

La
ortobiología es un nuevo procedimiento de la medicina regenerativa, la cual se
encarga de aplicar los conocimientos biológicos y biomecánicos; con esto se
indica que primero hay que tener un estudio y una base relacionada a las
funciones de cada uno de los componentes y células para saber qué beneficios y
consecuencias pueden ocurrir, y biomecánicos porque hay que tener un
conocimiento acerca del funcionamiento del aparato locomotor.
PLASMA RICO EN PLAQUETAS (PRP)
El PRP es plasma con una alta concentración de plaquetas que
contienen una gran cantidad de determinadas proteínas o factores de
crecimiento. Estas proteínas pueden iniciar y acelerar la reparación tisular en
tendones, músculos y ligamentos.
¿Para qué se utiliza el PRP?
Su aplicación está asociada a
lesiones tendinosas, desgarros musculares o roturas fibrilares, esguinces, etc.
Asimismo, se utiliza con éxito en lesiones cartilaginosas y en la
osteoartritis.
CÉLULAS MADRE
Las células madre son células mesenquimales, es
decir, capaces de diferenciarse en células de los diferentes tejidos, entre los
que se encuentran los del aparato locomotor (hueso, cartílago, tendón y
músculo).
¿Cómo funcionan las células madre?
Como las células madre son
multipotenciales, tienen la capacidad de diferenciarse en las células del
cartílago, tendón o músculo. El objetivo de cada tratamiento con células madre
es inyectar las células madre en las articulaciones para reparar o regenerar el
cartílago, o en los tendones o músculos para regenerarlos.
¿De dónde se obtienen las células madre y cómo se utilizan?
Pueden ser aisladas de una
amplia variedad de tejidos, incluyendo la médula ósea y el tejido adiposo que
son hoy los más utilizados para este propósito. En nuestro caso utilizamos el
tejido adiposo (grasa)La ventaja de usar las células madre del tejido adiposo
es que son una de las fuentes más ricas de células madre en el cuerpo .
Formación del Sistema Nervioso
(Neurulación)
La Neurulación es el proceso a través del cual se forma el Tubo Neural,
que da lugar a:
1. Sistema Nervioso Central: (Médula Espinal y Encéfalo)
1. Sistema Nervioso Central: (Médula Espinal y Encéfalo)
2. Cresta Neural,
(un grupo de células que migrarán por el resto del cuerpo y formarán Tejidos
Especializados como:
a. cartílagos y huesos
del cráneo,
b. neuronas, glía (soporte y protección de neuronas),
c. tejidos conectivos,
d. arcos faríngeos,
e. células del timo,
f. odontoblastos (primordio de los dientes), y
g. huesos del oído medio y de la mandíbula.
b. neuronas, glía (soporte y protección de neuronas),
c. tejidos conectivos,
d. arcos faríngeos,
e. células del timo,
f. odontoblastos (primordio de los dientes), y
g. huesos del oído medio y de la mandíbula.
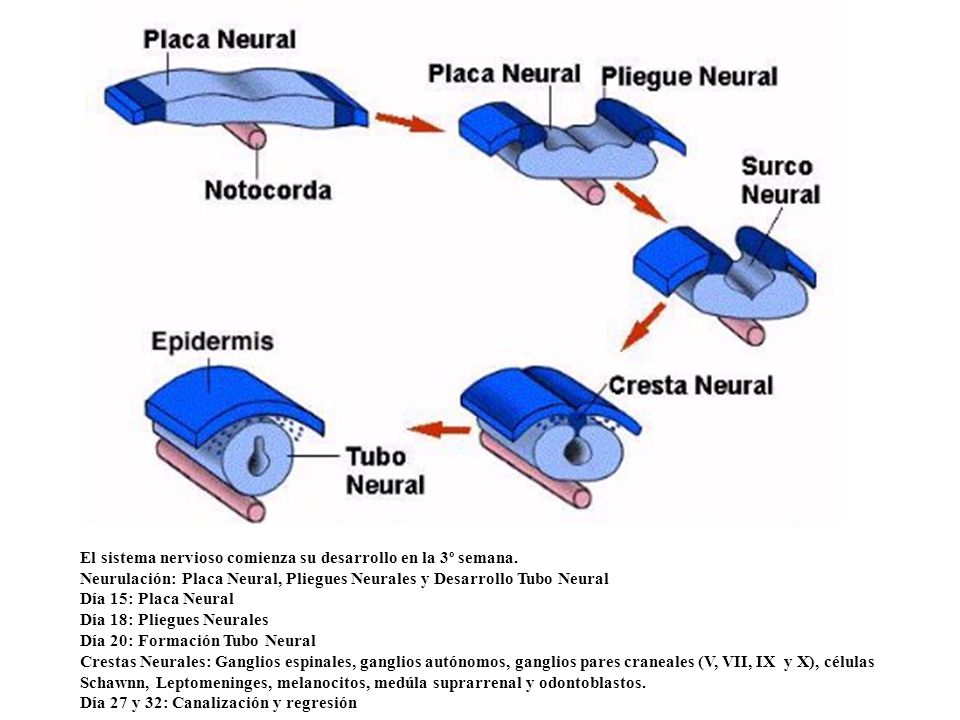
La neurulación o desarrollo del tubo neural comienza en la tercera
semana del embarazo, cuando se produce la gastrulación, a partir de la cual se
producirán todos los tejidos provenientes del ectodermo, mesodermo y endodermo.
En esta semana se distingue un esbozo del sistema nervioso, pero no será
hasta la sexta semana cuando aparezcan las estructuras básicas. además, a
diferencia del resto de sistemas y órganos, la organogénesis de este
sistema no termina en el primer trimestre, sino que continúa durante todo el embarazo
e incluso en los primeros meses de vida.
Finalmente el tubo neural se agranda por dentro y forma 3 vesículas
encefálicas primarias son:
- prosencéfalo (cerebro anterior),
- mesencéfalo (cerebro medio) y
- rombencéfalo (cerebro posterior).
En la quinta semana, las vesículas primarias se convierten en 5
vesículas secundarias:
- prosencéfalo -> telencéfalo y diencéfalo
- mesencéfalo -> se mantiene
- rombencéfalo -> metencéfalo y mielencéfalo.
También aparecen unas células muy peculiares, las células de la cresta
neural, situadas en el ángulo que forma el canal neural y el ectodermo que
migrarán más adelante a su lugar definitivo formando estructuras como los
ganglios raquídeos, parte del sistema nervioso autónomo o estructuras no
nerviosas como huesos de la cabeza.
Asimismo, es importante el espacio que se desarrolla entre
las paredes del tubo neural, el neurocele o canal neural, ya que a partir de él
se forman los ventrículos y el resto de cavidades del encéfalo, a través de los
cuales circula el líquido cefalorraquídeo.

Sistema Respiratorio
Aparecimiento del Primer esbozo de Aparato
Respiratorio.
1.Origen Endodérmico:
-Epitelio de
la laringe.
La Tráquea.
Los Bronquios y
Los Alveolos Pulmonares.
2. Origen Mesodérmico:
Estructuras Cartilaginosas.
Musculares.
Sistema Vascular.
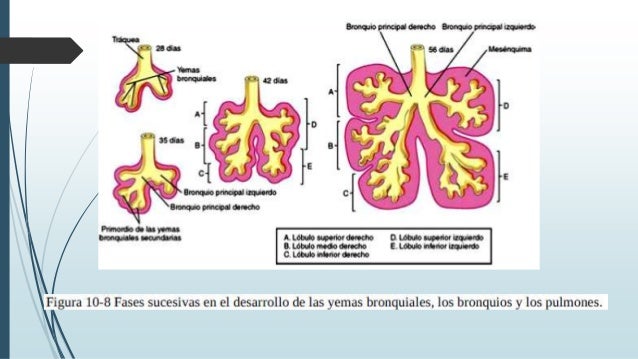
Aparecimiento del Primer esbozo de Aparato
Respiratorio en forma de evaginación de la pared ventral del intestino
anterior.
- El primordio respiratorio crece
caudalmente formando inicialmente la tráquea.
- De la tráquea surgen dos evaginaciones:
- Derecha: se divide ulteriormente en tres ramas.
- Izquierda: se divide en dos ramas.
- Sucesivas ramificaciones bronquiales prosiguen
hasta el sexto mes del desarrollo embrionario.
Aparecimiento del Primer esbozo de Aparato
Respiratorio en forma de evaginación de la pared ventral del intestino
anterior.
- El primordio respiratorio crece
caudalmente formando inicialmente la tráquea.
- De la tráquea surgen dos evaginaciones:
- Derecha: se divide ulteriormente en tres ramas.
- Izquierda: se divide en dos ramas.
- Sucesivas ramificaciones bronquiales prosiguen
hasta el sexto mes del desarrollo embrionario.

EMBRIOLOGÍA DEL SISTEMA
RESPIRATORIO
Los pulmones se cubren de una hoja
visceral (pleura visceral) que deriva del mesodermo.
La parte interna de la caja torácica
está tapizada por otra pleura parietal derivada de la hoja somática
mesodérmica. Las membranas pleuroperitoneales y otros
componentes musculares de la pared del cuerpo, conforman el diafragma.
Período Pseudoglandular (5ª –
16ª)
El epitelio cuboide bronquial
presenta un aspecto histológico que recuerda al de una glándula exocrina.
Se forma la «unidad respiratoria» o
«acino». los componentes de dicha unidad son: un bronquiolo terminal con 2 a 4 formaciones
tubulares y un conjunto de 6 a 7 sáculos.
Los acinos se ven progresivamente invadidos por capilares.Las células
epiteliales se van diferenciando.
Período Perinatal. Se producen los siguientes
cambios:
1.Movimientos respiratorios: por efectos
hormonales o mediadores químicos.
2. Reabsorción de líquido pulmonar:
a) alta concentración proteica.
b) cese de la bomba de sodio
epitelial.
c) insuflación del pulmón.
d) reabsorción a través de los linfáticos
pulmonares
e) reducción de la
presión intratorácica.

Período Posnatal (6ª - 8ª semana):
Tiene lugar un rápido desarrollo alveolar.
1) se produce una elongación de los bronquiolos
respiratorios.
2) los sáculos y los conductos transicionales se
transforman en conductos alveolares mediante un aumento de su longitud.
3) durante los años siguientes al nacimiento se van
produciendo cambios pulmonares hasta aproximadamente los 7 años.
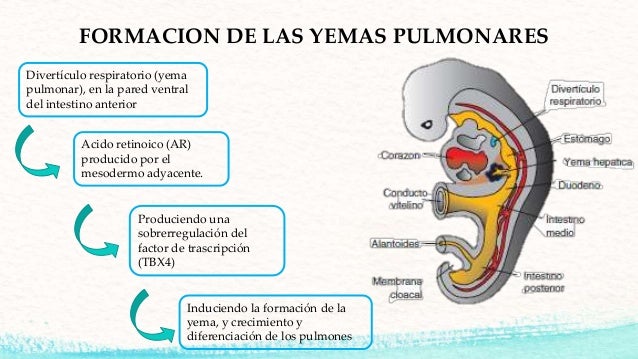
Sistema Digestivo
En la cuarta semana de gestación el embrión crece rápidamente,
este crecimiento favorece el aparecimiento de dos plegamientos.
Uno céfalo-caudal a consecuencia del cual, las
células que se convertirán en el corazón pasan de estar de la porción cefálica
a movilizarse hacia su cavidad definitiva.
Células endodérmicas en relación con el saco
vitelino se infiltran formándose el intestino primitivo y el conducto
denominado ónfalo mesentérico, conducto por el cual se comunicará el
saco vitelino con el intestino primitivo.
El pliegue lateral o transversal favorecerá
el cierre de cavidades. además los plegamientos del
embrión generan la formación de vasos sanguíneas
que irrigan completamente al embrión.
El intestino
primitivo está dividido en tres: intestino anterior, intestino medio e
intestino posterior. el intestino anterior, hacia cefálico, se divide en un
denominado intestino faríngeo, extendiéndose desde la membrana bucofaríngea en
relación con formación de estructuras respiratorias, y una porción caudal que
tendrá relación con estructuras digestivas.
Sistema Circulatorio
El
Sistema Circulatorio deriva del mesodermo.
La
implantación del embrión acelera los procesos de crecimiento y diferenciación,
restando eficiencia a la difusión y obligando a la formación del sistema
circulatorio cuyo principal objetivo es proporcionar los nutrientes al embrión
en desarrollo.
El
sistema circulatorio es el primero en aparecer y desarrollarse en el embrión.
Su órgano principal, el corazón, funciona antes que ningún otro.
Sistema circulatorio
en un embrión humano de 4 semanas.
Los grandes
vasos aparecen en un número par a derecha e izquierda, el
esquema representa solamente el lado derecho del embrión.

La disponibilidad de un sistema que posibilite el
transporte y el intercambio de sustancias de todas las células con su medio es
un requisito indispensable para construir un embrión que aumente de
tamaño y complejidad.
Las necesidades que debe cubrir el sistema
circulatorio varían durante el desarrollo embrionario de un individuo.
El sistema circulatorio se ve limitado por una
serie de imperativos fisiológicos, que condicionan su desarrollo.
Un embrión consta de células que necesitan
nutrientes y oxígeno o desechar metabolitos y co2. y todo esto es así
desde mucho antes de que se hayan desarrollado el intestino, los pulmones o el
riñón.
En las primeras etapas del desarrollo embrionario
el aporte de nutrientes hacia el embrión procede del saco vitelino y/o de la
placenta.
En el embrión la respiración no se realiza
por la vía pulmonar, sino que es canalizada a través de las membranas coriónica
y alantoidea.
Teniendo en cuenta que ambos procesos se llevan a
cabo por difusión, es necesario que estas membranas estén convenientemente vascularizadas.
esta circunstancia constituye una exigencia fisiológica y determina que los
primeros grandes vasos del embrión estén diseñados para irrigar a estas
estructuras extraembrionarias.
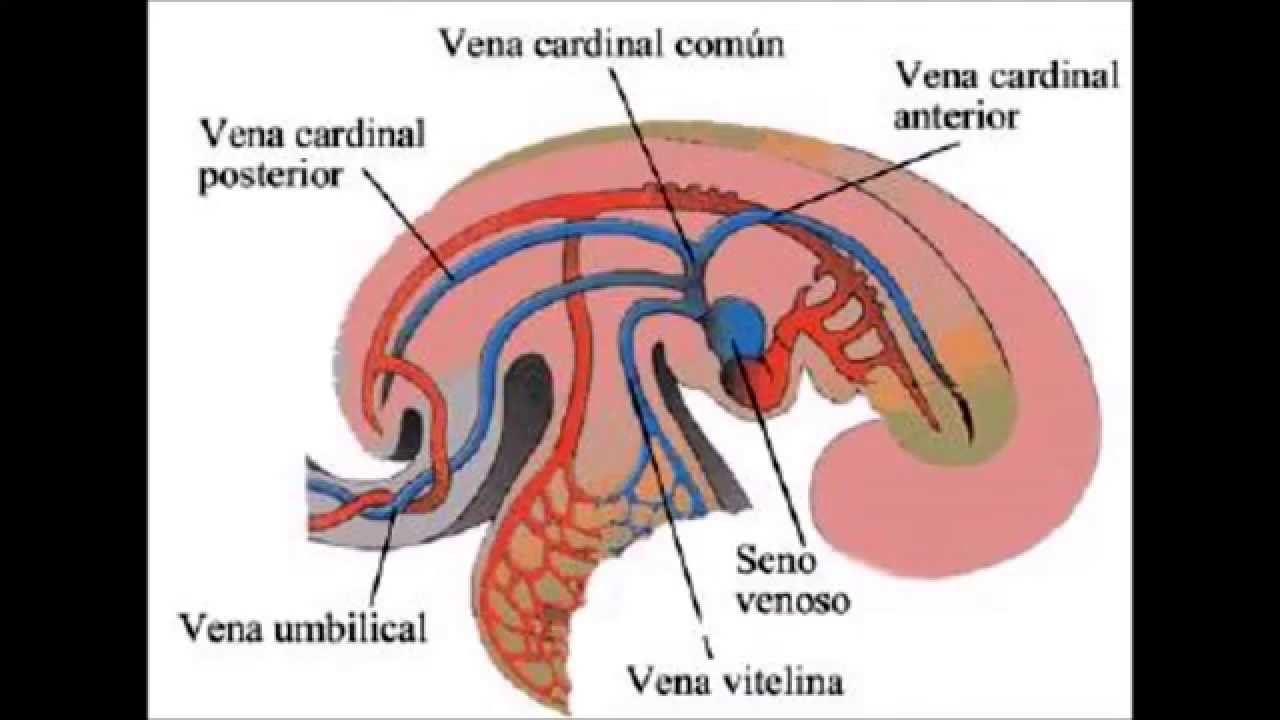
El embrión en una fase inicial, extiende sus vasos
sanguíneos hacia el vitelo. un poco más tarde, los vasos que abandonan
el primordio cardíaco forman seis pares de arcos aórticos que
confluyen en la aorta dorsal por encima de la faringe.
El embrión en una fase inicial, extiende sus vasos
sanguíneos hacia el vitelo.
Un poco más tarde, los vasos que abandonan
el primordio cardíaco forman seis pares de arcos aórticos que
confluyen en la aorta dorsal por encima de la faringe.
Un embrión debe ser entendido como un sistema
dinámico donde las necesidades cambian temporal y espacialmente.
Tanto el sistema circulatorio como el corazón
adquieren morfologías y, en consecuencia, funcionamientos diferentes durante el
desarrollo embrionario y el diseño debe ser el adecuado en cada uno de los
estadios.
Por ejemplo, durante el desarrollo fetal la parte
izquierda del corazón es deficitaria en oxígeno debido a que no existe retorno
pulmonar.

La formación temprana del sistema cardiovascular se correlaciona con la ausencia de una cantidad significactiva de vitelo en el óvulo y saco vitelino.
Al
final de la segunda semana, la nutrición embriónica se obtiene por
medio de la sangre materna mediante la difusión a través del trofoblasto.
las sustancias pasan entonces por el celoma extraembriónico y el saco
vitelino hacia el embrión. ya que éste inicia su desarrollo rápido en la
tercera semana, hay necesidad de vasos para que lleven sustancias nutritivas y
oxígeno hacia el embrión a partir de la circulación materna.
La formación de vasos sanguíneos inicia cuando las
células mesenquimatosas (angliobastos) , se unen y forman masas
aisladas y cordones llamados islotes sanguíneos.
Aparecen cavidades en estos islotes.
Las células mesenquimatosas se ordenan
alrededor de esas cavidades para dar origen al endotelio de los vasos
sanguíneos primitivos.
Los vasos endoteliales primitivos se funden para
configurar redes.
Los vasos se extienden hacia áreas adyacentes y se
unen con otros.
Las células sanguíneas y plasma se desarrollan
durante la tercera semana partiendo de células endoteliales de vasos en
las paredes del saco vitelino y de la alantoides.
La sangre inicia su formación en el embrión desde
la quinta semana.
El corazón primitivo es una estructura tubular que
se forma como un gran vaso sanguíneo a partir de las
células mesenquimatosas en el área cardiogénica .
Se desarrollan pares endocardiales de
vasos cardíacos y se funden para formar un corazón primitivo antes
del final de la tercera semana.
Al finalizar esta semana, los tubos cardíacos se
han unido a los vasos sanguíneos del embrión, al tallo o pedúnculo conector,
corion y saco vitelino, para formar un sistema cardiovascular primitivo. la
circulación de la sangre se inicia al final de la tercera semana cuando el
corazón tubular empieza a latir.
Aparato excretor
"Uno de los órganos más importantes de este sistema es el riñón, es un órgano en forma de frijol, aproximadamente del tamaño de nuestro puño, es un órgano retroperitoneal y se localiza más exactamente debajo de la caja torácica. Los riñones eliminan urea a través de sus unidades funcionales, la nefrona, cada nefrona posee una bola de capilares sanguíneos llamada glomérulo y una serie de túbulos llamados túbulos renales, la orina se produce a medida que pasa por los glomérulos y los túbulos, cada parte del túbulo tiene una función diferente para reabsorción y excreción. De los riñones la orina viaja a la vejigapor dos tubos de aproximadamente 8 a 10 pulgadas de largo, muy delgados llamados uréteres, alrededor de cada 10 a 15 segundo se vacían pequeñas cantidades de orina desde los uréteres y hacia la vejiga.La vejiga es un órgano muscular hueco en forma de globo que se encuentra ubicado sobre la pelvis, se encarga de almacenar la orina, puede llegar a almacenar hasta 16 onzas de orina de 2 a 5 horas, hasta que estemos listos para ir al baño a expulsarla."
La excreción consiste en eliminar de nuestro cuerpo los residuos producidos por la actividad celular. Estos residuos están disueltos en la sangre y son expulsados al exterior por el aparato excretor.
El sistema urinario empieza a desarrollarse a partir de 3 estructuras principales.
- · PRONEFROS: Formado por 7-10 grupos celulares que forman losnefrostomas, unidades vestigiales excretoras, al final de la cuarta semana el sistema pronéfrico desaparece dejando sus vestigios que son los nefrostomas.
- · MESONEFROS: Mientras se da la regresión del sistema pronéfrico, se forman los primeros túbulos excretores del mesonefro que se alargan formando una S y dando origen a una serie de capilares que formaran un glomérulo y alrededor del glomérulo los túbulos forman la capsula de Bowman y estas dos estructuras juntas forman el corpúsculo renal. El túbulo se inserta lateralmente en el conducto colector longitudinal llamadoconducto mesonéfrico o conducto de Wolff. Los túbulos caudales todavía se están desarrollando mientras que los craneales se degeneran y desaparecen en el aparato urogenital femenino.
- · METANEFROS: Estas unidades secretoras se desarrollan a partir delmesodermo metanéfrico a partir de él se desarrolla el riñón definitivo y elsistema colector y el sistema excretor.

El aparato respiratorio colabora en la
excreción, ya que mediante el intercambio de gases elimina el dióxido de
carbono.
SISTEMA COLECTOR:
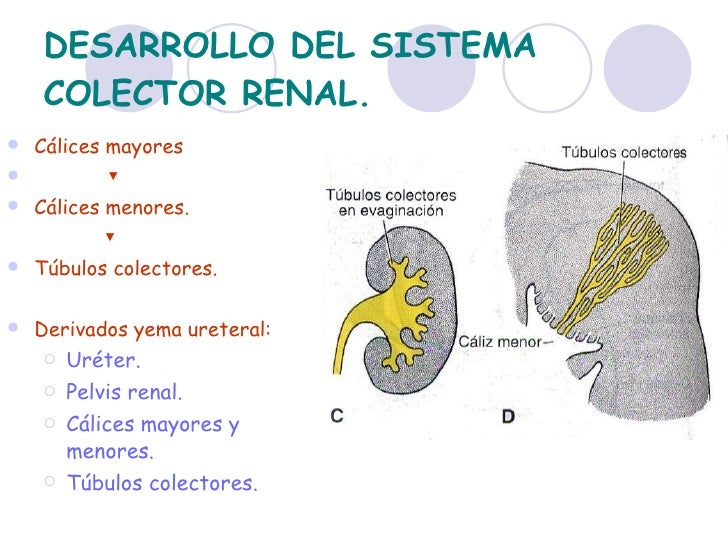
SISTEMA EXCRETOR:
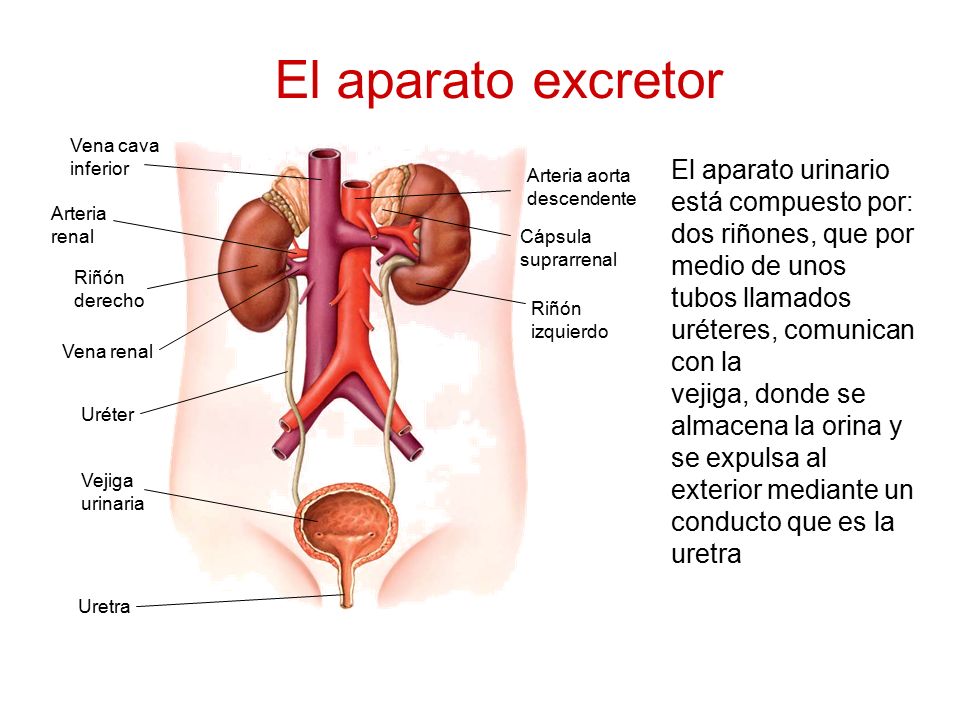
POSICIÓN DEL RIÑÓN:
FUNCIÓN DEL RIÑÓN:
SISTEMA COLECTOR:
Los conductos excretores del riñón se desarrollan a partir de una excrecencia del conducto mesonéfrico cerca de su entrada a la cloaca, llamado yema uretral. Más adelante la yema se dilata formando la pelvis renal primitiva que se divide en una parte craneal y otro caudal que serán lo cálices renales mayores. A medida que penetran el tejido metanéfrico se siguen dividiendo hasta llegar más o menos a 12 túbulos y este desarrollo continúa hasta finales del quinto mes llegando a la formación de los cálices menores. Posteriormente los túbulos colectores se alargan y convergen en el cáliz menor, formando allí la pirámide renal.
SISTEMA EXCRETOR:
Cada túbulo colector formado está cubierto por un casquete de tejido metanéfricoen su parte distal, estas células del casquete forman pequeñas vesículas llamadasvesículas renales y estas a su vez generan unos túbulos en forma de S.
Los capilares crecen en un extremo de la S y se diferencian en glomérulos, ambos, túbulos y glomérulos forman las nefronas o unidades excretoras, que es la unidad funcional del riñón, y el extremo proximal de cada nefrona desarrolla la cápsula de Bowman, mientras que el extremo distal y su alargamiento continúo da lugar a eltúbulo contorneado proximal, el asa de Henle y el túbulo contorneado distal.
Las nefronas se desarrollan hasta el momento del nacimiento, donde tenemos cerca de 1 millón de nefronas por riñón, al momento del nacimiento los riñones tienen una forma lobulada que con el tiempo desaparece debido al crecimiento de las nefronas, pero cabe resaltar que en este crecimiento no se aumenta su número o cantidad, simplemente su tamaño.
POSICIÓN DEL RIÑÓN:
Al comienzo del desarrollo urogenital el riñón se empieza a formar en la región pélvica en una posición caudal, pero a medida que se desarrolla pasa a la región abdominal en una posición más craneal debido al crecimiento y perdida de la curvatura corporal del feto.
FUNCIÓN DEL RIÑÓN:
El riñón definitivo formado a partir del metanefros pasa a ser un funcional en la semana 12, la orina pasa a la cavidad amniótica y se mezcla con el líquido amniótico, durante el periodo fetal el riñón no realiza su función de excreción de productos de desecho debido a que esta función es realizada por la placenta.
VEJIGA Y URETRA:
VEJIGA Y URETRA:
Aproximadamente de la 4ta a la 7ma semana la cloaca se divide anteriormente en elseno urogenital y posteriormente en el conducto anal, el seno urogenital se diferencia en 3 partes, la superior y además más grande es la vejiga urinaria.
Al principio la vejiga es continua con el alantoides pero cuando la luz del alantoides desaparece, el uraco se mantiene y se une al ápice de la vejiga y el ombligo que en el adulto es el ligamento umbilical medio. La siguiente parte del desarrollo origina la parte prostática y membranosa de la uretra, se origina es en el sexo masculino y es un conducto estrecho desarrollado a partir de la parte pélvica del seno urogenital.(imagen)
La última parte es la parte fálica del seno urogenital, esta aplanada en ambos extremos y a medida que el tubérculo genital crece, esta parte se extiende ventralmente, el desarrollo de la parte fálica del seno urogenital difiere ampliamente entre los dos sexos.
Durante la diferenciación de la cloaca, las partes caudales de los conductos mesonéfricos desaparecen absorbidos por las paredes de la vejiga y quedan como sus excrecencias los uréteres, los cuales entran a la vejiga por separado y como consecuencia del ascenso de los riñones los orificios uretrales se desplazan hacia una posición más craneal. Mientras tanto los orificios de los conductos mesonéfricos, en el sexo masculino se desplazan para introducirse en la uretra prostática para posteriormente pasar a ser los conductos eyaculadores.
El aparato excretor está formado por
el sistema o aparato urinario y por las glándulas sudoríparas.
El aparato urinario es el conjunto de
órganos que producen y excretan orina, el principal líquido de desecho del
organismo. El aparato urinario humano consta de los riñones, los uréteres, la
vejiga urinaria y la uretra.
La orina, que se forma en los riñones, se filtra a través de los uréteres, se acumula en la vejiga y es expulsada al exterior por la uretra.
La orina, que se forma en los riñones, se filtra a través de los uréteres, se acumula en la vejiga y es expulsada al exterior por la uretra.
Los riñones
Es la pareja de órganos cuya función es
la elaboración y la excreción de orina. En el ser humano, los
riñones se sitúan a cada lado de la columna vertebral, en la zona lumbar, y
están rodeados de tejido graso.
Tienen forma de judía. Miden unos 12 cm
de largo por 5 cm de ancho y 3 cm de grosor. Pesan alrededor de 150 gramos cada
uno.

En su interior se distinguen dos zonas:
la corteza, de color amarillento y situada en la periferia,
y la médula, zona más interna y rojiza. Un riñón es, en esencia, un
filtro que actúa como:
- Órgano regulador: mantiene en la
sangre una cantidad siempre igual de agua, sales y glucosa.
- Órgano depurador: el riñón extrae
de la sangre los productos nocivos, como la urea o el ácido úrico y los expulsa
al exterior.
a- Pelvis renal
Debajo de la corteza y la
médula se encuentra la pelvis renal, una cámara interior
subdividida que recolecta la orina y la dirige hacia el uréter. En ella entran
y salen, respectivamente, la arteria y la vena renal.
b- Nefrón
En la capa exterior se encuentran
diminutos filtros llamados nefronas, con una extensa red de
vasos sanguíneos que se encargan de filtrar la sangre y formar, gota a gota, la
orina que se reúne en la pelvis renal.
Cada nefrón consta de tres
partes principales:
- El glomérulo de Malpighi, una estructura
esférica formada por un ovillo de capilares sanguíneos de los cuales se toma el
líquido de la sangre a través de las paredes capilares porosas.
- La cápsula de Bowman, que tiene forma de
una copa hueca y presenta doble membrana. En su centro se encuentra el
glomérulo de Malpighi.
- Un túbulo renal, largo y
contorneado. Este fino tubo es la continuación de la cápsula de Bowman. Tiene
forma de asa y está rodeado de una red de capilares sanguíneos. Este tubulo se
divide en tres partes:
- El túbulo proximal: Es un conducto
sinuoso que sigue a la cápsula de Bowmann.
- El asa de Henle: Es un conducto
muy fino en forma de U.
- El túbulo distal: Desemboca al final en
uno de los túbulos renales o colectores que hay en la médula renal. Los túbulos
colectores desembocan finalmente en los cálices renales.
2.2- Vías excretoras
Conductos y cavidades que comunican los
riñones con el exterior.
a- Los uréteres
Los uréteres son dos largos tubos que
van desde la pelvis renal hasta la vejiga urinaria. Están constituidos por
fibra muscular lisa, y epitelio mucoso y terminaciones nerviosas. Son éstas las
que regulan el funcionamiento al ordenar contracciones que
impulsan la orina de forma continua y la hacen penetrar en la vejiga. Los
uréteres tienen terminaciones muy sensibles al dolor, de modo que cuando se
obstruyen, como ocurre en los cólicos nefríticos, se producen fuertes dolores.
b- La vejiga
La vejiga urinaria es el órgano hueco
en el que se almacena la orina formada en los riñones. La orina llega a la
vejiga procedente de los riñones por dos uréteres y se elimina hacia el
exterior a través de la uretra. La vejiga de la orina es un depósito elástico,
formado por fibra muscular lisa que tiene una capacidad que varía en torno a 1
litro, pero se tiene sensación de llenado ("ganas de orinar") desde
los 400 centímetros cúbicos.
c- La uretra
La uretra es el conducto a través del
cual se elimina la orina hacia el exterior. Es un tubo que parte de la zona
inferior de la vejiga y posee en su comienzo dos esfínteres o válvulas
musculares que controlan el paso de la orina. La uretra es diferente en cada
sexo, ya que en el varón interviene en la función reproductora. La uretra
femenina tiene una longitud de 3 a 4 cm y va desde la base de la vejiga al exterior,
terminando entre los dos labios menores, delante de la abertura vaginal. En la
uretra masculina, de 17 a 20 cm de longitud, se distinguen tres partes: porción
pélvica, rodeada por la próstata; porción membranosa y porción esponjosa. Esta
última corresponde al pene.
3- La orina y la micción
La orina es un líquido
de color amarillo claro que está compuesto por agua y otros elementos. El más
importante de estos elementos es la urea.
La orina se produce continuamente en el
riñón y llega a la vejiga intermitentemente, debido a los movimientos de los
uréteres. Unas válvulas impiden el retroceso de la orina desde la vejiga a los
uréteres. Cuando la vejiga está llena, se originan impulsos nerviosos que
producen el deseo consciente de orinar y de forma voluntaria se abre el
esfínter externo dando salida a la orina (micción).
La cantidad de orina que un adulto
normal elimina, por término medio, cada 24 horas, es de 1,5 litros (un
litro y medio). Este volumen varía con la cantidad de líquido y alimento
ingerido así como con las pérdidas por vómitos o a través de la piel por la
sudoración.
La orina está compuesta de: 95 %
de agua, 2 % de sales minerales, cloruros, fosfatos, sulfatos, sales
amoniacales, 3% de sustancias orgánicas, urea, ácido úrico, ácido
hipúrico y creatinina.
3.1- ¿Cómo se forma la orina?
La sangre entra en los riñones para que las nefronas vayan retirando los productos de desecho y el exceso de sales y formen la orina. Los procesos que se llevan a cabo en la nefrona para la formación de orina son:
La sangre entra en los riñones para que las nefronas vayan retirando los productos de desecho y el exceso de sales y formen la orina. Los procesos que se llevan a cabo en la nefrona para la formación de orina son:
Filtración
La sangre que llega a la nefrona es filtrada en la cápsula de Bowman, pudiendo ser filtradas todas las sustancias de pequeño tamaño excepto las grandes moléculas y las células. El producto resultante tiene una composición parecida al plasma sanguíneo y contiene muchas sustancias aprovechables por el organismo.
La sangre que llega a la nefrona es filtrada en la cápsula de Bowman, pudiendo ser filtradas todas las sustancias de pequeño tamaño excepto las grandes moléculas y las células. El producto resultante tiene una composición parecida al plasma sanguíneo y contiene muchas sustancias aprovechables por el organismo.
Reabsorción
El filtrado va pasando por los tubos de la nefrona donde se produce una reabsorción selectiva de las sustancias aprovechables que pasan a la sangre a veces por transporte activo con gasto de energía . También se reabsorbe gran cantidad de agua.
El filtrado va pasando por los tubos de la nefrona donde se produce una reabsorción selectiva de las sustancias aprovechables que pasan a la sangre a veces por transporte activo con gasto de energía . También se reabsorbe gran cantidad de agua.
Secreción
Algunas sustancias que no se han filtrado o se han reabsorbido equivocadamente son secretadas desde los capilares sanguíneos que rodean a la nefrona al interior de los tubos de esta, obteniéndose por último la orina final.
Algunas sustancias que no se han filtrado o se han reabsorbido equivocadamente son secretadas desde los capilares sanguíneos que rodean a la nefrona al interior de los tubos de esta, obteniéndose por último la orina final.
La orina ya formada va saliendo de la
nefrona hacia el comienzo del uréter por donde baja a la cloaca o a la vejiga
urinaria.
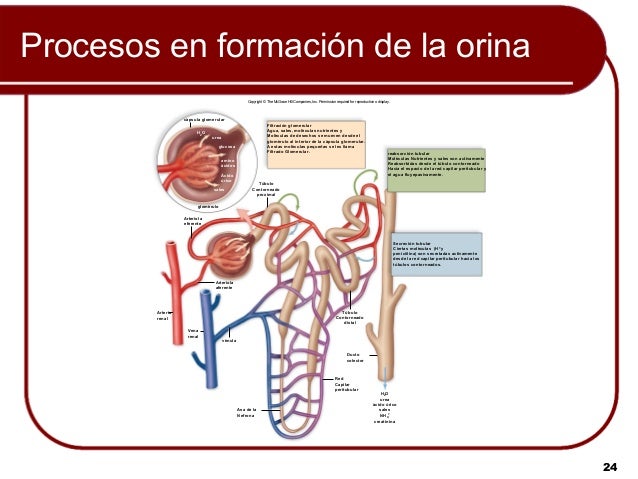
4- Algunas enfermedades del aparato
urinario
La especialidad médica que estudia el
aparato urinario se llama urología y el médico especialista es
el urólogo.
4.1- Insuficiencia renal
Es la incapacidad de los riñones para filtrar y depurar la sangre. Como consecuencia de esta alteración se acumulan en el organismo productos tóxicos que deberían ser eliminados o, por el contrario, se eliminan sustancias que deberían conservarse, como proteínas o glóbulos rojos. Cuando la insuficiencia renal es grave, se recurre al riñón artificial. Este aparato no cura al riñón enfermo, simplemente sustituye sus funciones. El proceso de filtrado o depuración que realiza el riñón artificial se conoce con el nombre de diálisis. Por término medio, este aparato logra purificar toda la sangre en un plazo aproximado de seis horas.
Es la incapacidad de los riñones para filtrar y depurar la sangre. Como consecuencia de esta alteración se acumulan en el organismo productos tóxicos que deberían ser eliminados o, por el contrario, se eliminan sustancias que deberían conservarse, como proteínas o glóbulos rojos. Cuando la insuficiencia renal es grave, se recurre al riñón artificial. Este aparato no cura al riñón enfermo, simplemente sustituye sus funciones. El proceso de filtrado o depuración que realiza el riñón artificial se conoce con el nombre de diálisis. Por término medio, este aparato logra purificar toda la sangre en un plazo aproximado de seis horas.
4.2- Cistitis
Inflamación de la vejiga urinaria, normalmente debida a una infección bacteriana originada en la uretra, vagina o, en casos más complicados, en los riñones. La cistitis también puede deberse a la irritación causada por los depósitos cristalinos de la orina, o a cualquier condición o anormalidad urológica que obstaculice el funcionamiento normal de la vejiga.
Entre los síntomas se pueden citar la micción dolorosa o dificultosa, la necesidad urgente de orinar y, en algunos casos, orina turbia o con sangre. El tratamiento consiste básicamente en tomar antibióticos, beber grandes cantidades de líquidos e inhibir la bacteria acidificando la orina (por ejemplo, tomándose infusiones de hojas de arándano).
Inflamación de la vejiga urinaria, normalmente debida a una infección bacteriana originada en la uretra, vagina o, en casos más complicados, en los riñones. La cistitis también puede deberse a la irritación causada por los depósitos cristalinos de la orina, o a cualquier condición o anormalidad urológica que obstaculice el funcionamiento normal de la vejiga.
Entre los síntomas se pueden citar la micción dolorosa o dificultosa, la necesidad urgente de orinar y, en algunos casos, orina turbia o con sangre. El tratamiento consiste básicamente en tomar antibióticos, beber grandes cantidades de líquidos e inhibir la bacteria acidificando la orina (por ejemplo, tomándose infusiones de hojas de arándano).
4.3- Nefritis
Denominación común para los procesos inflamatorios del riñón. Existen varias formas de nefritis. La aparición de algunas formas se manifiesta por cansancio, pérdida de apetito, inflamación de la cara, dolor abdominal o en el costado, y disminución del volumen de orina, que además es más oscura de lo habitual. Otra característica principal es la presencia en la orina, en el examen microscópico, de albúmina (lo que se denomina albuminuria).
Denominación común para los procesos inflamatorios del riñón. Existen varias formas de nefritis. La aparición de algunas formas se manifiesta por cansancio, pérdida de apetito, inflamación de la cara, dolor abdominal o en el costado, y disminución del volumen de orina, que además es más oscura de lo habitual. Otra característica principal es la presencia en la orina, en el examen microscópico, de albúmina (lo que se denomina albuminuria).
Es mucho más frecuente en la infancia y
adolescencia que en la edad adulta. Las formas que son más frecuentes en niños,
son también las de mejor pronóstico en general, las que tienen una mejor
recuperación. Otras formas pueden terminar en nefritis crónica, la cual produce
una lesión progresiva y destructiva del riñón.
4.4- Cálculos renales
Son más conocidos como piedras en el riñón. Se pueden formar en el riñón o en la pelvis renal por depósitos de cristales presentes en la orina. Los cálculos pueden producir hemorragia, infección secundaria u obstrucción. Cuando su tamaño es pequeño, tienden a descender por el uréter hacia la vejiga asociados con un dolor muy intenso. El dolor cólico producido por los cálculos requiere tratamiento con analgésicos potentes, y puede aparecer de forma súbita tras el ejercicio muscular.
Son más conocidos como piedras en el riñón. Se pueden formar en el riñón o en la pelvis renal por depósitos de cristales presentes en la orina. Los cálculos pueden producir hemorragia, infección secundaria u obstrucción. Cuando su tamaño es pequeño, tienden a descender por el uréter hacia la vejiga asociados con un dolor muy intenso. El dolor cólico producido por los cálculos requiere tratamiento con analgésicos potentes, y puede aparecer de forma súbita tras el ejercicio muscular.
Una vez que el cálculo alcanza la
vejiga, es posible que sea expulsado por la orina de forma inadvertida,
desapareciendo el dolor. Si el cálculo es demasiado grande para ser expulsado,
es necesario recurrir a la cirugía o a la litotricia, procedimiento que utiliza
ondas de choque generadas por un aparato localizado fuera del organismo,
para desintegrar los cálculos.
4.5- La uremia
Es la intoxicación producida por la acumulación en la sangre de los productos de desecho que suelen ser eliminados por el riñón. Aparece en la fase final de las enfermedades crónicas del riñón. La generalización de los trasplantes de riñón han supuesto un gran avance para estos pacientes.
Es la intoxicación producida por la acumulación en la sangre de los productos de desecho que suelen ser eliminados por el riñón. Aparece en la fase final de las enfermedades crónicas del riñón. La generalización de los trasplantes de riñón han supuesto un gran avance para estos pacientes.
5- Las glándulas sudoríparas
Las glándulas sudoríparas son las
encargadas de excretar el sudor. El sudor contiene agua, sales minerales y un
poco de urea. No obstante, la misión excretora de las glándulas es secundaria.
Su principal función es la de regular la temperatura corporal mediante la
evaporación del agua expulsada. En algunos momentos se puede perder hasta 1
litro de agua por hora. Las glándulas sudoríparas están repartidas por toda la
piel, pero son más numerosas en la cabeza, axilas y palmas de las manos.
ht
DESARROLLO
DE LOS ORGANOS DE LOS SENTIDOS
•El desarrollo progresivo del tacto, la vista , el
gusto, el olfato y el oído a lo largo del desarrollo embrionario y fetal cumple
con una doble función:
•1. Modelar su sistema nervioso central, y
•2. Preparar al nuevo ser para disponer
de organos que le ayuden a relacionarse e integrarse a su vida en el
mundo exterior.
Tacto
•Es
el primer sentido que se pone en funcionamiento. la primera señal de
sensibilidad aparece en torno a las siete semanas alrededor de la boca.
•Durante
las dos semanas siguientes, la piel que es sensible a la estimulación táctil se
extiende al resto de la cara, a las palmas de las manos y al tórax.
•En
la semana 11ª las plantas de sus pies y los genitales muestran una gran
sensibilidad, y ya a las 12 semanas se amplía a toda la superficie del cuerpo,
salvo la coronilla y la espalda.
•A
partir del sexto o séptimo mes, abre la boca y se chupa el dedo. a
través de pequeños receptores cutáneos el feto también detecta las leves
variaciones de temperatura que se producen en el útero y las vibraciones
Vista
•En
torno a los 22 días de gestación en el embrión se hacen evidentes los surcos
que darán lugar a la estructura ocular. son dos concavidades muy separadas que
se van aproximando muy rápidamente.
•En
el octavo mes es sensible a la luz, las pupilas se contraen o
dilatan dependiendo de la intensidad lumínica, y también abre los ojos cuando
está despierto y los cierra cuando está dormido.
•Hacia
el cuarto mes los ojos son sensibles a la luz. puede apreciar si un fuerte estímulo
luminoso (el sol) atraviesa la pared uterina y el líquido amniótico y reacciona
cambiando de posición para protegerse de la luz.

Gusto
•Los
sentidos del gusto y el olfato están muy relacionados y en el cerebro se
perciben de una forma muy similar.
•A
la sexta semana ya es reconocible la lengua y sus papilas gustativas comienzan
a formarse una semana después.
•A
partir del tercer mes el sistema gustativo está activo y en torno al cuarto mes
es capaz de distinguir diferentes sabores.
•Desde
el tercer mes de gestación, el sentido del gusto esta desarrollado. si percibe
un sabor dulce en el líquido amniótico, acelera los movimientos de deglución,
mientras que, si este contiene un sabor amargo o que le resulta desagradable,
deja de tragar en el acto.
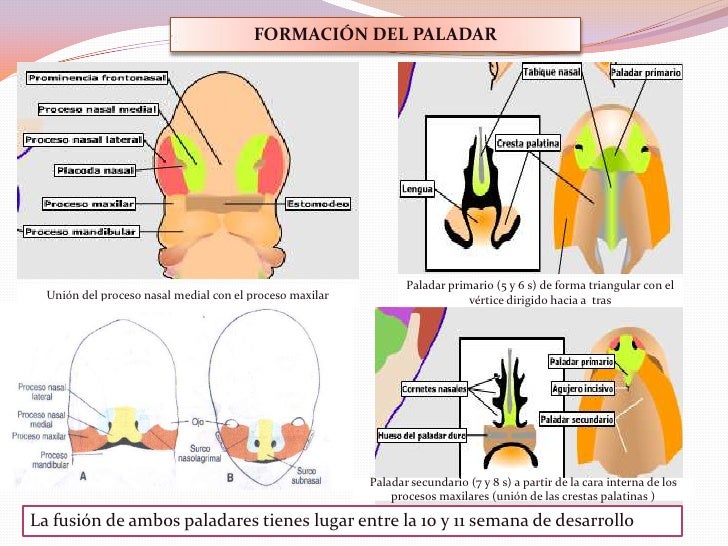
Olfato
•A
las cinco semanas comienza a aparecer un esbozo de nariz en
la cabeza del embrión. se trata
de las plácodas nasales, que se sitúan en la región
frontal, pero habrá que esperar algunas semanas para que este órgano empiece a
hacerse visible.
•La
depresión superficial que forman estos puntos da lugar a las fosas nasales;
alrededor de ellas se forman unas elevaciones de tejido en forma de herradura
que se transformarán en las aletas de la nariz.
•Entre
las semanas 6ª y 7ª se desarrollan la punta, la cresta nasal y parte del
tabique, como resultado de lo cual, a la 8ª semana, la nariz está
diferenciada. a las siete semanas los nervios y la membrana olfatoria también
están formados y a partir de la 12ª semana la nariz está preparada para
percibir todo tipo de olores.
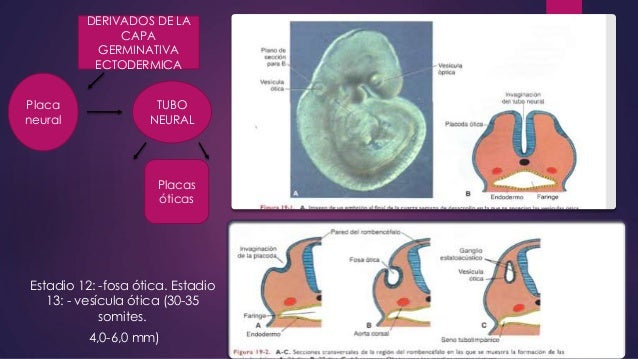
Oído
•El
oído comienza a formarse durante las primeras semanas en los arcos
braquiales, poco a poco, los pabellones auditivos se desplazan hasta
su localización definitiva.
•Al
final del sexto mes, el oído ya está completamente formado. sin embargo, el
feto puede percibir sonidos procedentes de la madre o del exterior desde la
16ª, y reacciona a ellos acelerando el ritmo cardíaco y la frecuencia de los
movimientos corporales.
•El
oído interno está totalmente desarrollado a partir de la mitad del embarazo y
el feto puede percibir los latidos del corazón de su madre, los ruidos
intestinales durante la digestión, el paso de la sangre a través del cordón
umbilical.

VIDEO
v
Bibliografías
http://desarrolloembrionariosistemarinario.blogspot.com/
https://es.slideshare.net/danbar1908/embriologia-sistema-respiratorio
https://es.slideshare.net/fredyliestrada/embriologa-del-sistema-cardiovascular-47119983
https://es.slideshare.net/Elcentinela1422/embriologia-aparato-digestivo

